Molecular Characteristics and Potent Immunomodulatory Activity of Fasciola hepatica Cystatin
Article information

Abstract
Cystatin, a cysteine protease inhibitor found in many parasites, plays important roles in immune evasion. This study analyzed the molecular characteristics of a cystatin from Fasciola hepatica (FhCystatin) and expressed recombinant FhCystatin (rFhcystatin) to investigate the immune modulatory effects on lipopolysaccharide-induced proliferation, migration, cytokine secretion, nitric oxide (NO) production, and apoptosis in mouse macrophages. The FhCystatin gene encoded 116 amino acids and contained a conserved cystatin-like domain. rFhCystatin significantly inhibited the activity of cathepsin B. rFhCystatin bound to the surface of mouse RAW264.7 cells, significantly inhibited cell proliferation and promoted apoptosis. Moreover, rFhCystatin inhibited the expression of cellular nitric oxide, interleukin-6, and tumor necrosis factor-α, and promoted the expression of transforming growth factor-β and interleukin-10. These results showed that FhCystatin played an important role in regulating the activity of mouse macrophages. Our findings provide new insights into mechanisms underlying the immune evasion and contribute to the exploration of potential targets for the development of new drug to control F. hepatica infection.
INTRODUCTION
Fascioliasis is an important zoonotic disease mainly caused by Fasciola hepatica, The disease leads to reduce growth rates and severely affects the production of livestock products such as meat, wool and milk from infected animals, posing a serious threat to animal industry [1]. In addition, F. hepatica is an important foodborne parasite that infects humans through raw vegetables and water contaminated with metacercariae [2]. To date, it is estimated that at least 2.4 million people are infected and 200 million of people are at risk [3]. Hence, fascioliasis is recognized as an important foodborne neglected tropical disease (NTD) by World Health Organization (WHO) [4].
When encysted metacercariae enter host’s intestine, the newly excysted juveniles (NEJ) can rapidly pass through the gut wall and migrate to the liver. To cross host’s physical barriers, NEJ can secrete excretory-secretory products (ESPs) such as cathepsin L-like and B-like cysteine proteases, cystatins, and Kunitz-type inhibitors, which contribute to the parasite’s tissue invasion and evasion from the host immune responses [5,6]. Among them, immunomodulation of host cells is a survival strategy often adopted by the parasite. In order to survive in host for a long time, F. hepatica can employ ESPs to induce cell apoptosis and degrade host immune molecules, thereby suppressing the activity of immune cells [7–9]. Cystatin is one of cysteine protease inhibitors with the activity of papain-like protein [10]. As a virulence factor, cystatin is proposed to be involved in regulating the humoral and cellular immune responses during the infection of parasites. Currently, it has been proved that parasite cystatins can act on a variety of cells, among which macrophages are one of its target cells [11].
Although cystatin has been identified in several parasites, the molecular characteristics and mechanism of its immune evasion have not been fully elucidated to date. With the in-depth study of F. hepatica genome and proteomics, more proteins with immunomodulatory effects have been identified. Several studies have shown that some proteins (e.g., F. hepatica Stefins-1, F. hepatica Stefins-2 and F. hepatica Stefins-3) in F. hepatica ESPs can inhibit mammalian cathepsin, displaying regulatory effects on host immune cells [12]. As a potent immunomodulatory molecule in F. hepatica, the roles of FhCystatin in host’s immune responses have not been explored in depth. Here, the molecular characteristics and enzymatic activity of a FhCystatin were characterized. Its regulatory roles on macrophage function were explored, which may provide new insights into the molecular mechanism of immune evasion exploited by F. hepatica.
MATERIALS AND METHODS
Ethical approval
All experiments were carried out in accordance with the guidelines issued by the Ethical Committee of Shihezi University (No. A20180126).
Parasites, animals, and cells
The worms of F. hepatica were collected from naturally infected sheep in Xinjiang Uygur Autonomous Region, China. After morphological identification, the species identity was confirmed as F. hepatica based on PCR and sequencing of the second internal transcribed spacer (ITS2) of ribosomal DNA. The parasites were stored at −80°C. Six-week-old BALB/c mice were purchased from the Experimental Animal Center of Shihezi University. Mouse macrophages RAW264.7 was cultured in DMEM (Gibco, New York, New York, USA) medium supplemented with 10% fetal bovine serum (Gibco) at 37°C incubator with 5% CO2.
Primer design
Based on the genomic and transcriptomic database of F. hepatica (AY647146.1), primers were designed to amplify the FhCystatin gene, forward primer: (5′-CCGGAATTCATGTTGCGCATACTACTTG-3′) and reverse primer: (5′-GCGGCCGCTCAAGTGCAGGATACCCGAGTC-3′). For the expression of FhCystatin protein, the restriction sites EcoR I and Not I were introduced in the upstream and downstream primers, respectively.
Analysis of molecular characterization of FhCystatin protein
The total RNA of F. hepatica was extracted using Trizol (Sigma, St. Louis, Missouri, USA), and the RNA was reverse transcribed into cDNA using a reverse transcription kit (Takara, Nojihigashi, Japan), and the cDNA was used as a template to amplify the FhCystatin gene. The amplified product was cloned into pMD19-T vector (Takara) for sequencing. The molecular characteristic of deduced amino acid sequence, such as molecular weight, isoelectric point, signal peptide, and transmembrane region, were analyzed using the online software ProParam (http://web.expasy.org/protparam/), ProtScale (http://expasy.org/tools/protscale.htmL), Signal P (http://www.cbs.dtu.dk/services/SignalP/), and TMHMM (http://www.cbs.dtu.dk/services/TMHMM/), respectively. Sequence comparison was performed using Clustal software and the 3D structure of the FhCystatin protein was predicted by SWISS-MODEL (https://www.swissmodel.expasy.org/).
Expression and purification of recombinant FhCystatin
The FhCystatin gene was subcloned into the pPIC9K vector (Invitrogen, San Diego, California, USA) to generate the pPIC9K-FhCystatin recombinant plasmid, and positive clones were screened using PCR and restriction digestion. Then, pPIC9K-FhCystatin was linearized with Sal I and electrotransformed into P. pastoris GS115 strain (Invitrogen) to screen the recombinant yeast strain. The recombinant yeast was cultured in BMGY medium and continuously induced with methanol (1%) for 5 day. Subsequently, the cells were precipitated by centrifugation and the culture supernatant was collected for SDS-PAGE analysis. Homogeneity of rFhCystatin was examined by 15% reducing SDS-PAGE. Western blot analysis was performed using Fh-infected sheep serum as primary antibody (dilution ratio 1:1,000) and horseradish peroxidase-conjugated rabbit anti-sheep IgG (Abcam, Cambridge, UK) as secondary antibody (dilution ratio 1:3,000). rFhCystatin was purified using Ni-NTA chromatography (Merck, Darmstadt, Germany) according to the manufacturer’s instructions.
Determination of the biological activity of rFhCystatin
Cathepsin B (Solarbio, Beijing, China) and Z-Arg-Arg-AMC (Sigma) were used as the target enzyme and substrate for the analysis of rFhCystatin protein activity, respectively. Briefly, cathepsin B (20 ng/reaction) was activated in buffer (0.1 M sodium acetate, 1 mM DTT, and 2 mM EDTA) at room temperature for 5 min. Then, rFhCystatin (ranging from 1 to 8 μM) was added and pre-incubated at room temperature for 20 min. Subsequently, the substrate Z-Arg-Arg-AMC was added and incubated at 37°C for 30 min. The relative activity (IC50 value) was calculated by detecting the fluorescence released from the hydrolysis of the substrate at 355 nm and 460 nm, respectively, using a multifunctional enzyme marker (Synergy2, Biotek, Burlington, Vermont, USA). In addition, rFhCystatin was incubated with buffers at different temperatures (25, 30, 37, 42, 45, and 50°C) and different pH (4–8) for 20 min, and the fluorescence values were detected to determine the optimum temperature and pH for rFhCystatin activity.
Interaction between rFhCystatin and RAW264.7 cells
Mouse RAW264.7 cells (105 cells per ml) were cultured and inoculated into confocal culture dishes (Solarbio). After the incubation with rFhCystatin (20 μg/ml) at 37°C for 2 h, these cells were fixed in 4% paraformaldehyde for 30 min at room temperature and washed 3 times with PBS (5 min/time). Then, cells were incubated with mouse anti-FhCystatin antibody (dilution 1:200) at 4°C for 12 h. Cells were washed 3 times with PBS and incubated with Cy3-conjugated goat anti-mouse IgG (Sigma) as secondary antibody (dilution 1: 500) at 37°C for 1 h. After that, cells were incubated with Hoechst 33342 (Sigma-Aldrich, St. Louis, Missouri, USA) to stain cell nuclei and observed using a laser confocal microscope (Nikon C2, Nikon, Tokyo, Japan).
Effects of rFhCystatin on macrophage proliferation and migration
The effects of rFhCystatin on the viability of RAW264.7 cells were detected using Cell counting Kit-8 (Beyotime, Haimen, China). Mouse RAW264.7 cells (105 cells per ml) were inoculated into 96-well tissue culture plates and incubated with different concentrations (1.0, 2.5, and 5.0 μg/ml) of rFhCystatin at 37°C for 48 h. After incubation, 10 μl of CCK-8 reagent was added to each well and incubated for 4 h protected from light. The cell survival index was calculated according to the OD values. Furthermore, the migratory roles of rFhCystatin on RAW 264.7 cells were determined as previously described [13].
Effects of rFhCystatin on the production of nitric oxide and cytokines in macrophage
Mouse RAW264.7 cells (106 cells per ml) were cultured on 12-well plates and subsequently co-treated with different doses of rFhCystatin (1.0, 2.5, and 5.0 μg/ml), lipopolysaccharide (LPS) (1 μg/ml) and PBS was used alone as control. After 24 h, the NO content in the cells was measured using a Nitric Oxide Assay Kit (mLbio, Shanghai, China). The concentrations of interleukin-6 (IL-6), interleukin-10 (IL-10), tumor necrosis factor-α (TNF-α) and transforming growth factor-β (TGF-β) cytokines in the supernatant of RAW264.7 cell culture were measured by enzyme-linked immunoassay kit (Merck).
Effects of rFhCystatin on apoptosis in macrophages
RAW264.7 cells were cultured on 6-well plates at a density of 106 cells/ml. Cells were treated with different concentrations of rFhCystatin (1.0, 2.5, and 5.0 μg/ml) for 24 h in a 37°C incubator with 5% CO2, and then treated with Annexin V-EGFP/PI Apoptosis Detection Kit (Beyotime) by flow cytometry (FACSAria III, Becton Dickinson, Franklin Lakes, New Jersey, USA) to assess and quantify apoptosis rates of RAW264.7 cells.
Statistical analysis of data
All data analyses were performed using GraphPad Premier 8.0 software (GraphPad Prism, San Diego, California, USA). Statistical analysis was performed using one-way ANOVA. The data are expressed as mean±standard deviation. Differences between test groups were considered significant if P-value was less than 0.05 (*P<0.05, **P<0.01, ***P<0.001, and ****P<0.0001).
RESULTS
As shown in Supplementary Figs. S1 and S2, the open reading frame of FhCystatin gene was 351 bp in length and encoded 116 amino acids. The deduced amino acid sequence contains a signal peptide located between amino acids 20 and 21. The N-glycosylation site and transmembrane region were not found. The sequence contained a conserved cystatin-like domain (Fig. 1A, B), the N-terminal region of conserved glycine residues and a central Q-X-V-X-G motif (Supplementary Fig. S3). FhCystatin consists of the α-helix (35.3%), β-fold (5.2%), and irregularly curled region (33.6%), which shared 92.2% identities with type I Cystatin (AFV53480.1) of F. gigantica at the protein level. The phylogenetic analysis showed that Fhcystatin and F. gigantica cystatin-1 were most closely related to each other and distantly related to cystatin from other species (Supplementary Fig. S4).
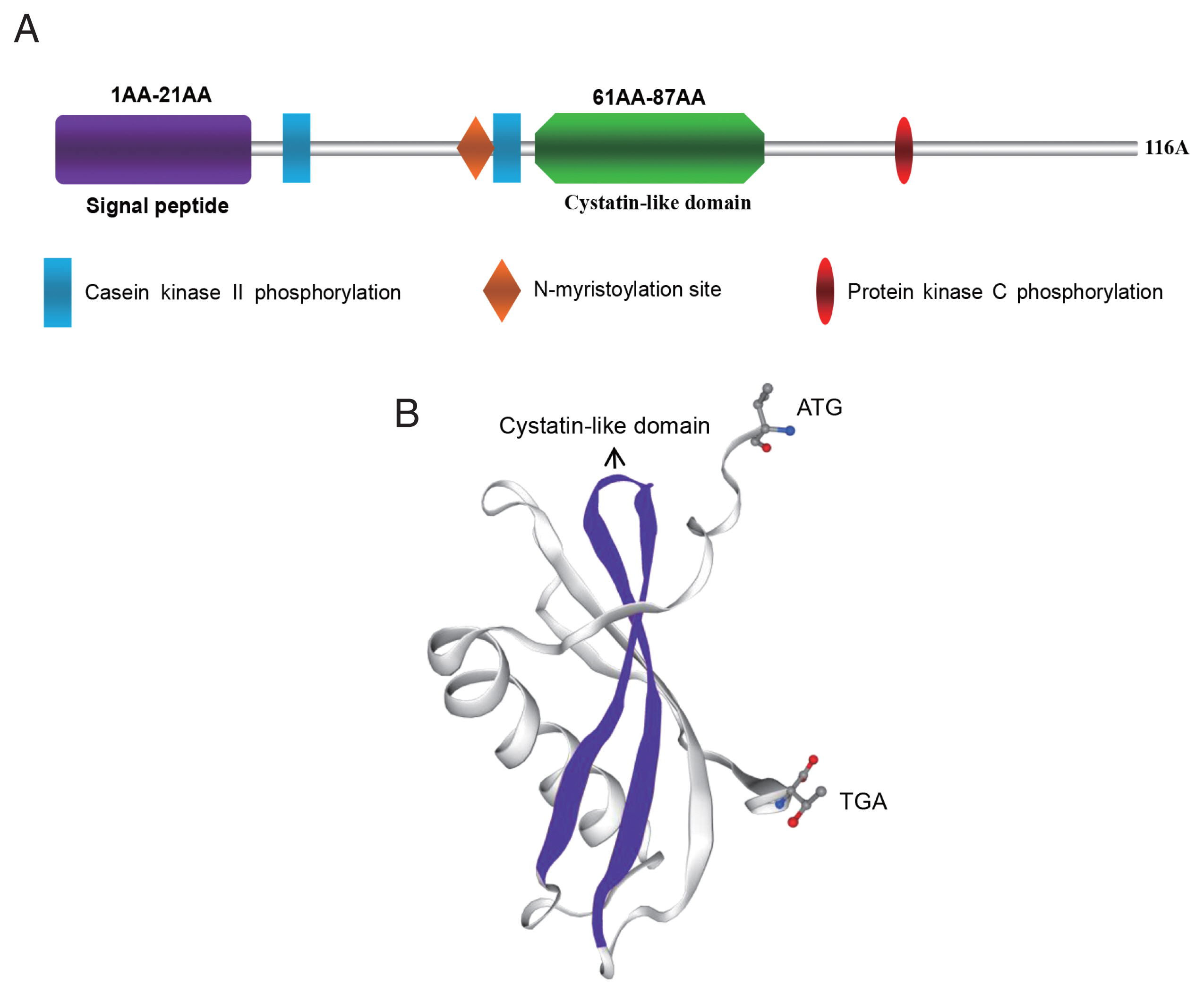
Molecular characterization of FhCystatin of Fasciola hepatica. (A) Schematic structural composition of the main domains of FhCystatin. (B) 3D structure model of FhCystatin (SWISS-MODEL).
The pPIC9K-FhCystatin plasmid was constructed, and positive clones were screened using PCR and restriction digestion (Supplementary Fig. S5). As shown in Fig. 2, recombinant FhCystatin (rFhCystatin) was expressed in P. pastoris, with an expected molecular weight at 12.47 kDa. Western blot confirmed that the rFhCystatin protein was recognized specifically by sheep serum infected with F. hepatica.
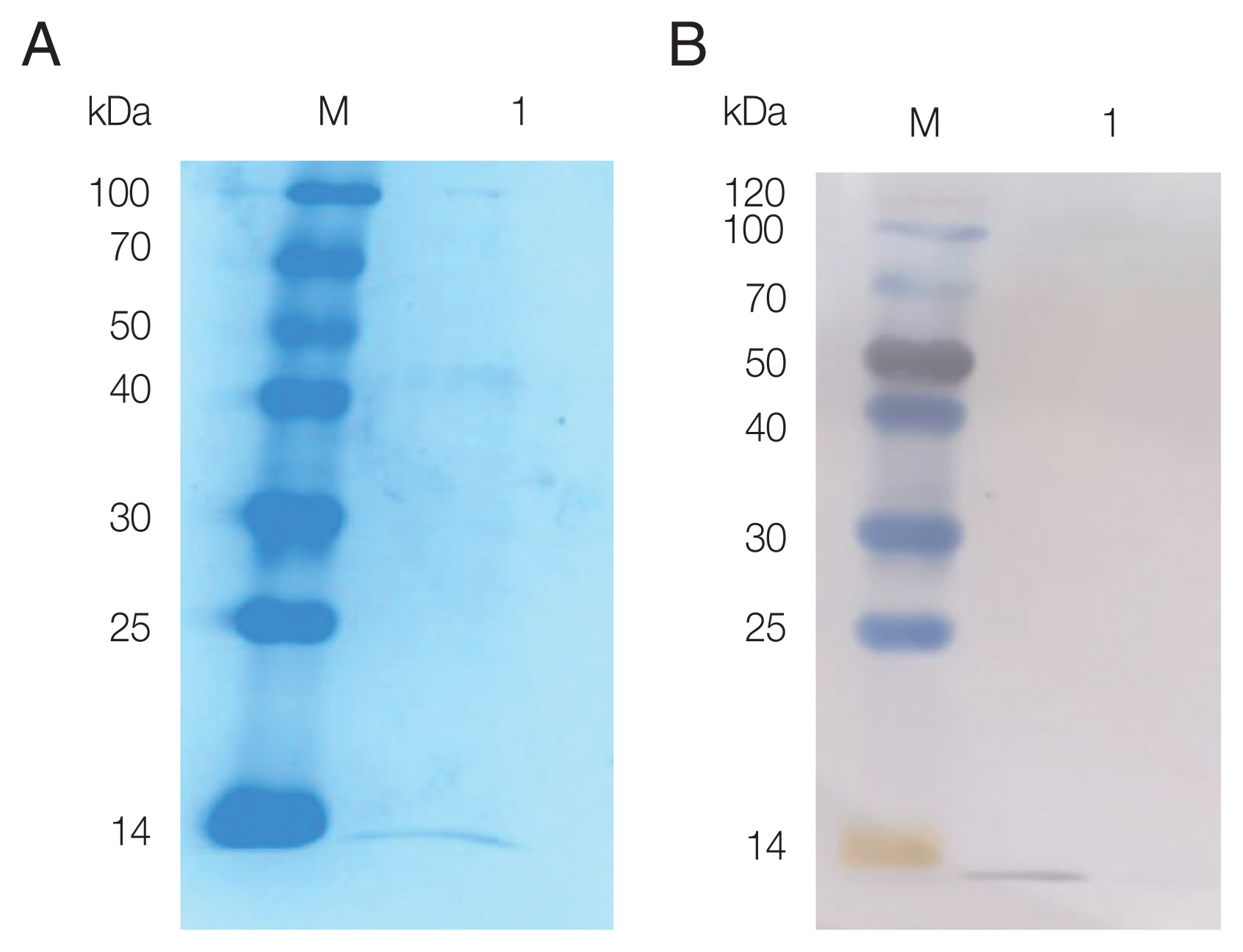
SDS-PAGE and western blot analysis of rFhCystatin expressed by P. pastoris. (A) Protein was resolved on 15% acrylamide gels and stained with Coomassie brilliant blue R250. Lane M: Standard molecular weight marker; Lane 1: Purified rFhCystatin. (B) The interest protein was run under non-reducing conditions, and visualized by immunodetection using specific antibodies by enhanced chemiluminescence. Lane M: Standard molecular weight marker; Lane 1: Purified rFhCystatin.
RFhCystatin inhibited cathepsin B activity in a dose-dependent manner. The inhibitory activity of rFhCystatin varied greatly at different pH and temperature; the maximum inhibitory activity was observed at pH 6 and at 37°C (Fig. 3). As shown in Fig. 4, rFhCystatin was bound to the surface of RAW264.7 cells. In contrast, control group failed to show the fluorescence of the red dye Cy3 on the surface of RAW264.7 cells.
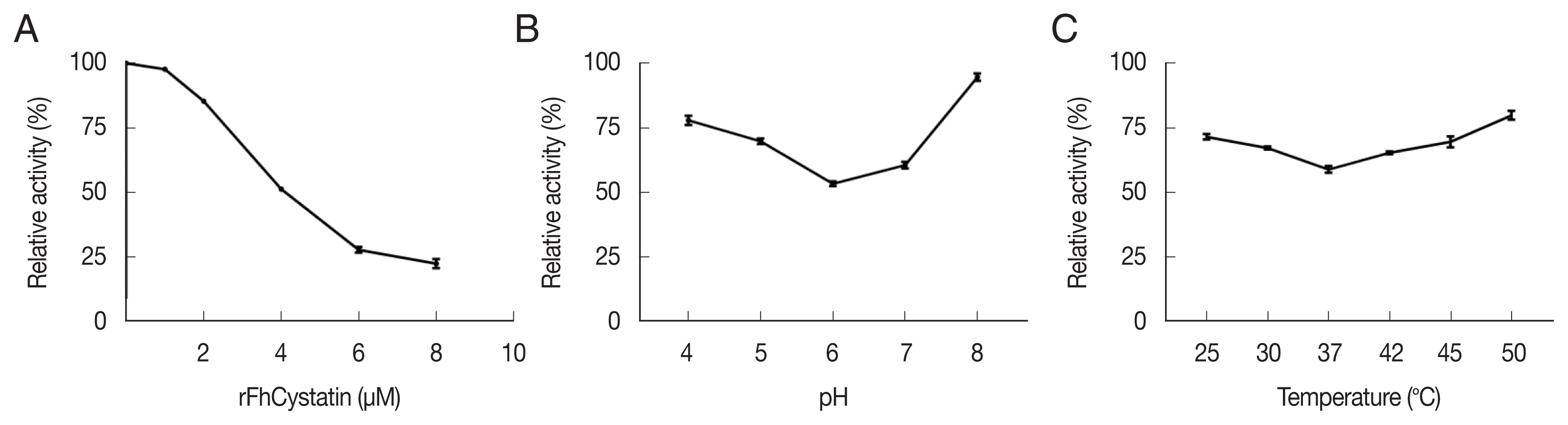
Determination of biological activity of rFhCystatin of Fasciola hepatica. (A) Different concentrations of rFhCystatin were incubated with cysteine proteases at room temperature for 20 min, after which residual enzyme activity was assayed. (B) rFhCystatin was incubated with cysteine proteases in different pH for 20 min at room temperature, after which residual enzyme activity was assayed. (C) rFhCystatin was incubated with cysteine proteases at different temperatures for 20 min at indicated, after which residual enzyme activity was assayed.
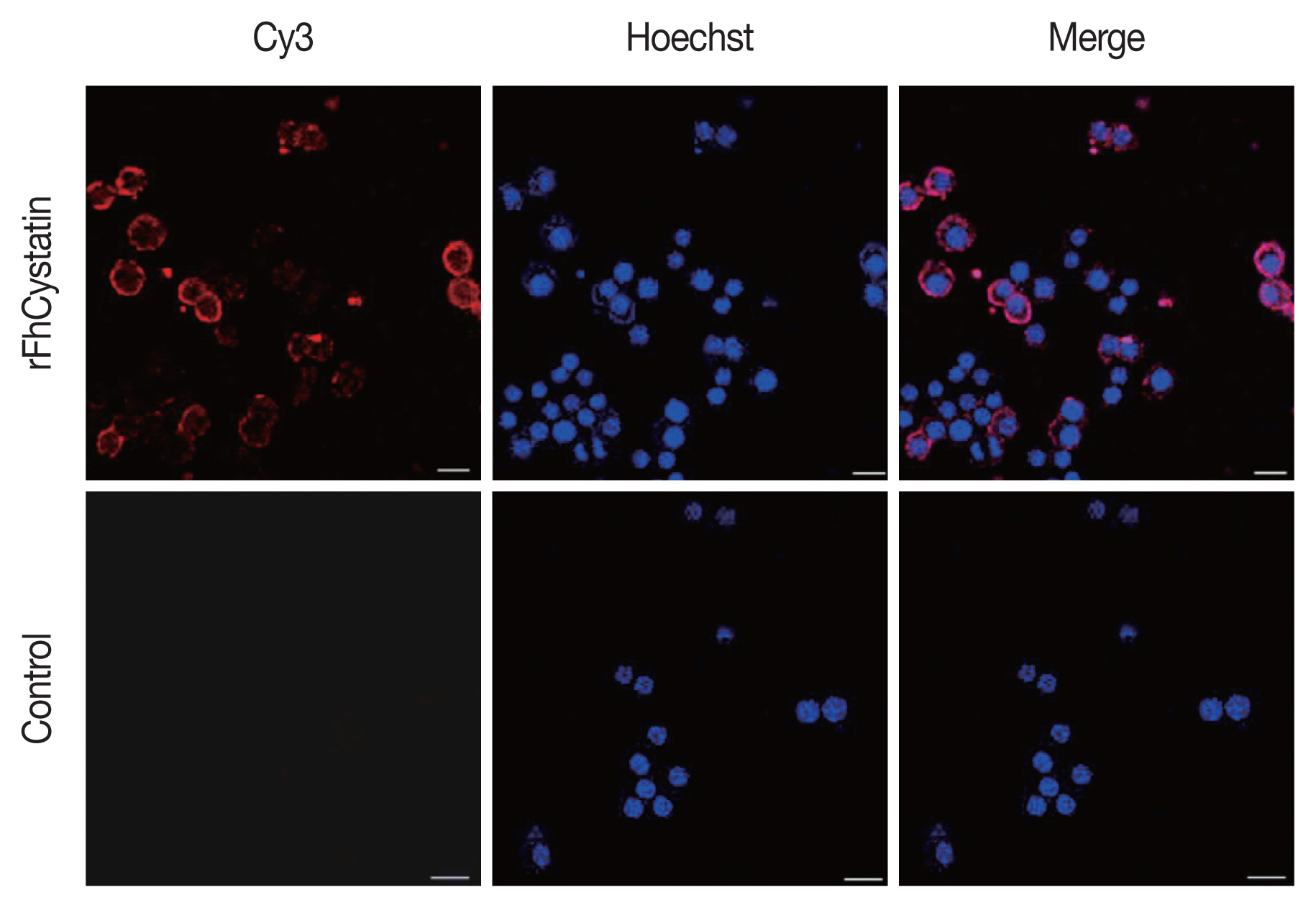
Fasciola hepatica-derived rFhCystatin bound to the surface of Raw264.7 cells. Visualization of rFhCystatin protein attachment to Raw264.7 cells surface was carried out by incubation of Raw264.7 cells treated or untreated with rFhCystatin with mouse anti-rFhCystatin antibody. Hoechst (blue) and Cy3-conjugated secondary antibody (red) were used to stain host cell nuclei and rFhCystatin, respectively. Surface staining was detected in rFhCystatin-treated cells. No staining was detectable in untreated cells. Scale-bars=15 μm.
Fig. 5 demonstrated that rFhCystatin (5 μg/ml) had a significant inhibitory effect on the proliferation of RAW264.7 cells compared with PBS control and low concentration of rFhCystatin (P<0.05). Moreover, rFhCystatin promoted the mobility of LPS-activated RAW264.7 cells (P<0.05) and displayed a dose-dependent effect.
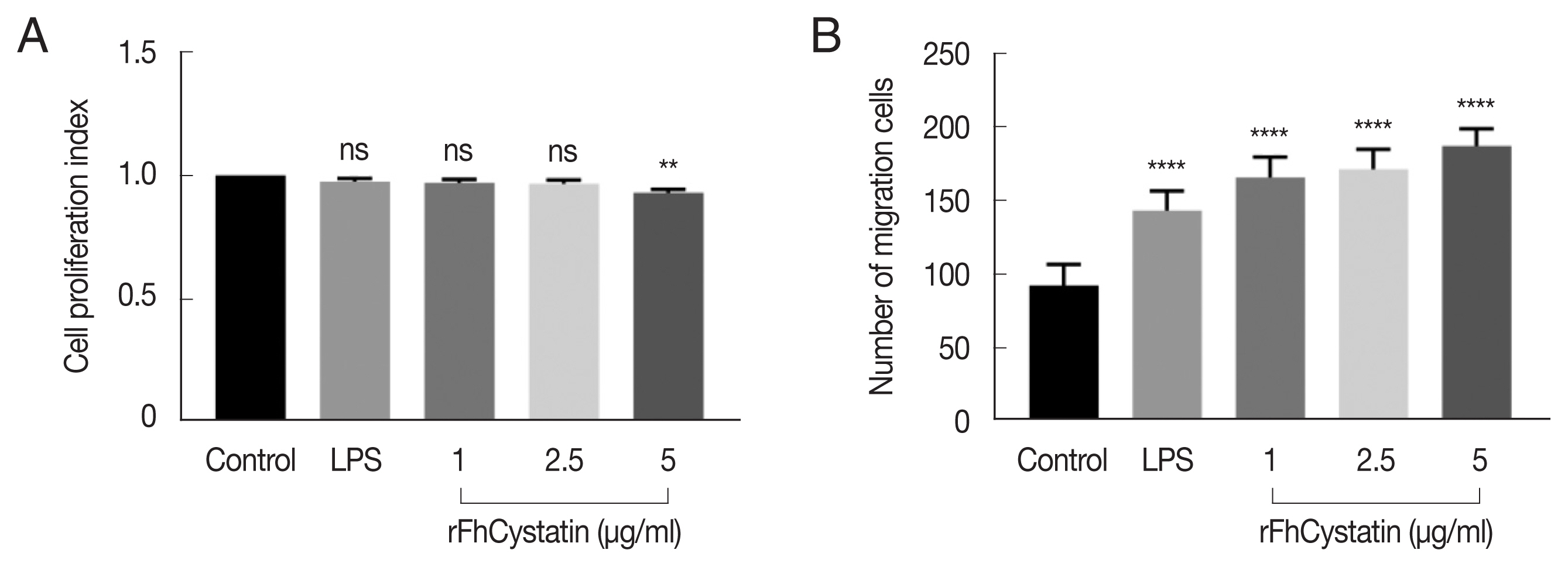
The effects of rFhCystatin on the proliferation and migration of RAW264.7 cells. (A) rFhCystatin inhibited RAW264.7 cell proliferation. RAW264.7 cells were sham-treated with control buffer or with different concentrations of rFhCystatin. Proliferation was determined using CCK-8 assay. (B) rFhCystatin inhibited RAW264.7 cell migration. Cells were sham-treated with control buffer or with different doses of rFhCystatin. The cell migration number was determined. Graphs represent the means±standard deviations of data from 3 independent biological replicates. ns, not-significant. **P<0.01, ****P<0.0001.
We observed effect of rFhCystatin on NO production. As shown in Fig. 6, rFhCystatin inhibited NO production in a dose-dependent manner in RAW264.7 cells compared to positive control (LPS) (P<0.05). Cytokine assays revealed that rFhCystatin inhibited the secretion of inflammatory factors IL-6 (P<0.01) and TNF-α (P<0.01) in RAW264.7 cells, while rFhCystatin significantly promoted expression of anti-inflammatory factors IL-10 (P<0.01) and TGF-β (P<0.01) secretion.
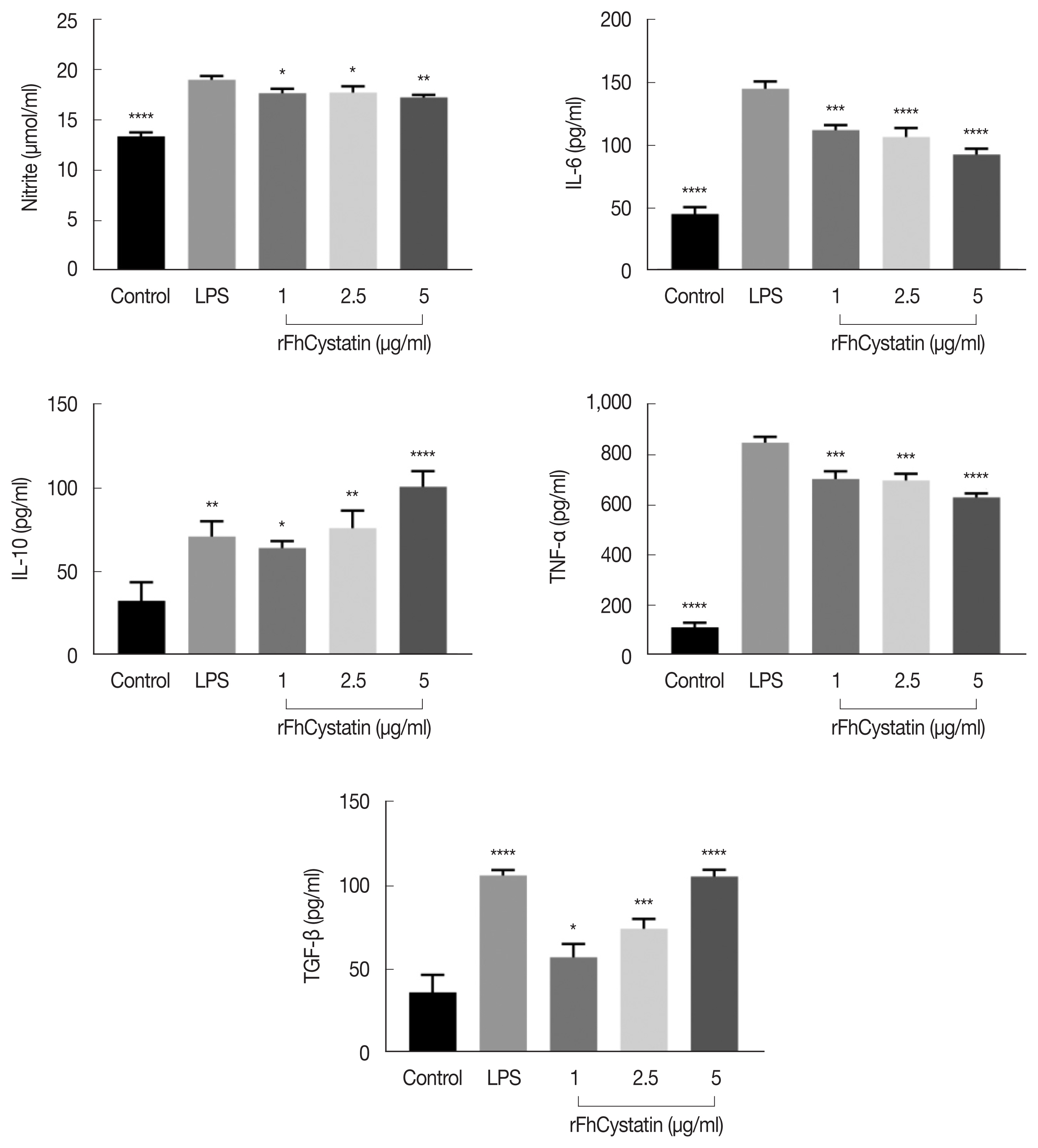
The effects of rFhCystatin on NO and cytokine production in RAW264.7 cells. RAW264.7 cells were incubated for 24 h in the presence or absence of different concentrations of rFhCystatin. The levels of NO and cytokine concentration in the supernatant of cultured cells were quantified by ELISA. Graphs represent the means±standard deviations of data from 3 independent biological replicates. ns, not-significant. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
As shown in Fig. 7, Anenxin V-FITC/PI double staining apoptosis assay revealed that rFhCystatin significantly induced apoptosis in RAW264.7 cells (P<0.01) and showed a dose-dependent effects.
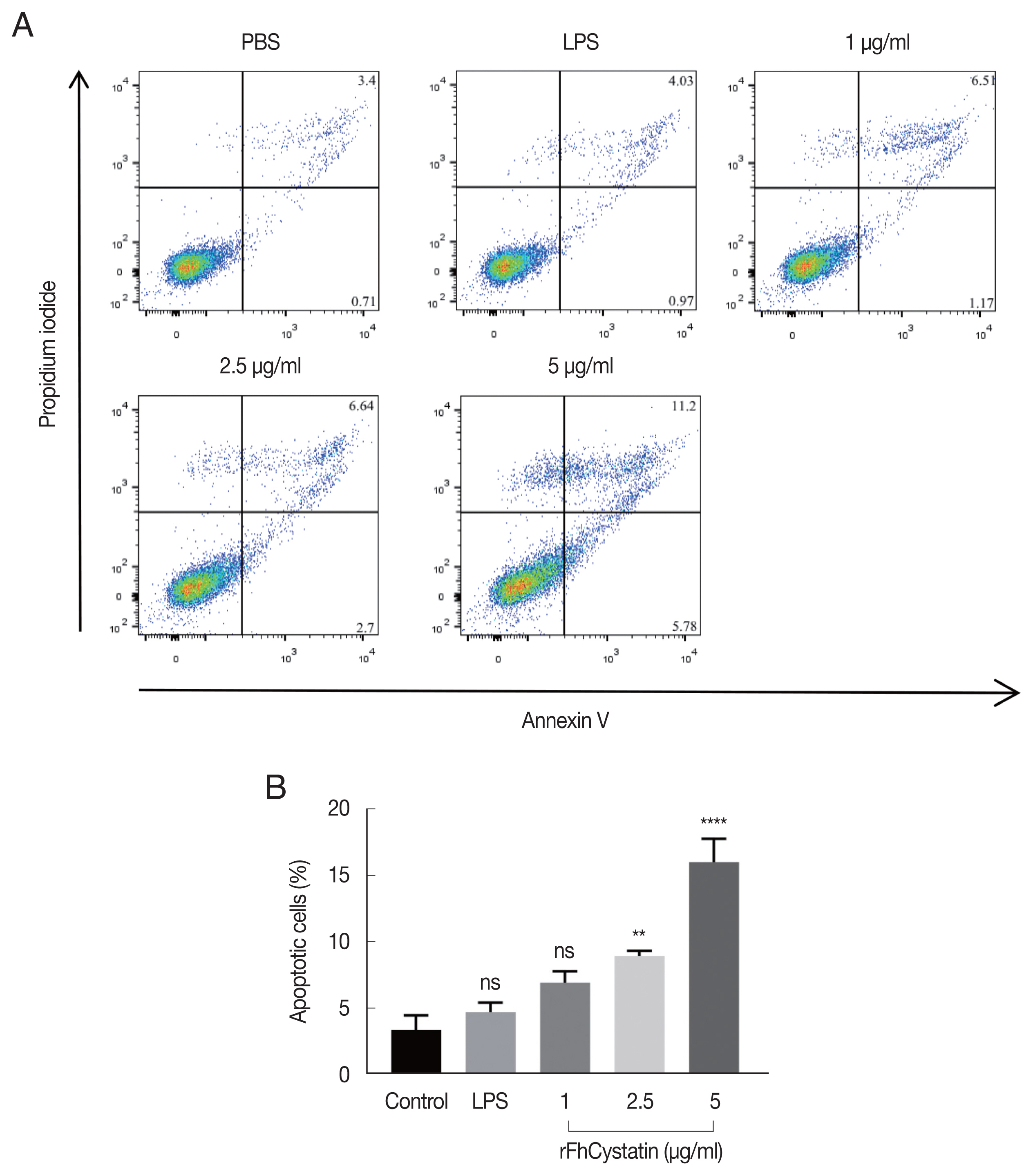
rFhCystatin induced apoptosis of RAW264.7 cells. Apoptotic cells were determined by Annexin V/PI staining with flow cytometry analysis. (A) Dot plot showing death of RAW264.7 cells in response to exposure to rFhCystatin. (B) Apoptotic cells (Annexin V+/PI−) were plotted and compared with percentage of cell population. Graphs represent the means±standard deviations of data from 3 independent biological replicates. ns, not-significant. **P<0.001, ****P<0.0001.
DISCUSSION
Several studies have revealed that excretory-secretory products of F. hepatica (FhESPs) can induce apoptosis in eosinophils and peritoneal macrophages, and modify host immune responses, resulting in host immune suppression [14]. However, the molecules mechanism of immune modulation still remains unclear, which constitutes a serious obstacle to developing protective vaccines in ruminants [15]. A few molecules of FhESPs were known to meditate the parasite’s invasion [16] and most of which are poorly understood in biological functions. This shortcoming has hindered the development of new drugs and vaccines against F. hepatica infection.
Many studies have shown that immune evasion is one of the most important strategies of long-term survival in the host for parasites. Previous studies have found that parasites can achieve immune evasion through a variety of mechanisms (e.g., intracellular parasitism, antigenic mutation, and immune regulation) [17–19]. During parasite infection, cystatin displayed modulatory effects on host immune responses [20]. In this study, FhCystatin gene was cloned and the molecular characteristics of encoded protein were analyzed. The results showed that FhCystatin contained an N-terminal conserved glycine residue and a central Q-X-V-X-G motif, confirming that it is similar in molecular structure to other parasite cystatins and belongs to the cystatin family.
Several studies have revealed that cysteine protease inhibitors (cystatins) are one of the components in the ESPs of F. hepatica [21]. However, to date, the knowledge of expression profiles of different cystatins in F. hepatica was limited. Considering that cysteine proteases are involved in cytophagy, intracellular clearance and antigen processing in the antigen-presenting cell (APC), cystatin produced by the parasite might inhibit the host’s cysteine protease activity, interfering with the function of APC, and modulate the host’s immune response [22,23]. In Echinococcus weissei, cystatin was found to promote the production of IL-10 and reduce the production of NO and TNF-α in macrophages, thus exerted anti-inflammatory effects [24]. The similar immunomodulatory effects of cystatin were also found in Schistosoma twigginum, mouse intestinal parasitic nematodes, and Schistosoma huaji [21,25,26]. In Schistosoma japonicum, cystatin stimulated the production of IL-10, but repressed the production of NO, IL-6, and TNF-α in macrophages, thereby played an important role while immune evasion [17].
Macrophages are one of important phagocytic cells that play crucial roles in the process of antigen processing. Macrophages regulated other immune cells through secretion of a variety of cytokines [27]. This study showed that FhCystatin bound to RAW264.7 cells and significantly inhibited the proliferation. Furthermore, FhCystatin suppressed the secretion of inflammatory factors IL-6 and TNF-α produced by RAW264.7 cells, while it significantly promoted the secretion of anti-inflammatory factors IL-10 and TGF-β. These findings suggested that FhCystatin exerted strong immunosuppressive effects. Therefore, it is speculated that the FhCystatin may be an important immunomodulatory molecule for F. hepatica to achieve immune evasion. The detailed mechanism by which FhCystatin modulates host immune responses requires further study.
It has been reported that macrophages produced oxidative molecules such as NO and reactive oxygen to attack and damage the worm during the course of infection [28]. We showed that the FhCystatin reduced NO production and the proliferation of RAW264.7 cells in a dose-dependent manner. However, the molecular mechanism by which FhCystatin protein suppresses NO production and proliferation of macrophages needs to be investigated.
Several studies have shown that parasitic ESPs induced the apoptosis of host cells [13]. The peritoneal leukocytes in sheep infected with F. hepatica underwent apoptosis to help larvae survive during peritoneal migration [29]. A previous study reported that liver flukes might reduce the release of inflammatory factors by inducing apoptosis of goat peripheral blood mononuclear cells (PBMCs) [30]. In turn, apoptotic cells could produce anti-inflammatory cytokines IL-10 and TGF-β to suppress the host immune responses [31–33]. Therefore, it can be inferred that macrophages apoptosis induced by FhCystatin may also be one of the survival strategies for F. hepatica to evade host immune response.
This study for the first time demonstrated that FhCystatin inhibited not only the proliferation of mouse macrophages, reduced the production of NO, IL-6, and TNF-α, and promoted the production of IL-10 and TGF-β, but also the apoptosis of mouse macrophages. These findings suggested that FhCystatin plays important immunomodulatory effects on the immune function of macrophages. FhCystain also plays multiple roles in the immunological pathogenesis of F. hepatica, which provide new insights into the mechanisms of immune evasion in F. hepatica. Our study may contribute to the identification of potential targets for the development of new drug to control F. hepatica infection.
Supplementary Information
ACKNOWLEDGMENTS
This work was supported by International Science & Technology Cooperation Program of XPCC (No. 2016AH006), Xinjiang Autonomous Region graduate innovation project (No. XJ2020 G085) and Grant from National Key Research and Development Program (No. 2017YFD0501202). We thank the staff who provided the technical assistance for this study.
Notes
The authors declare that they have no conflict of interest.