Paragonimus and Paragonimiasis in Vietnam: an Update
Article information
Abstract
Paragonimiasis is a food-borne parasitic zoonosis caused by infection with lung flukes of the genus Paragonimus. In Vietnam, research on Paragonimus and paragonimiasis has been conducted in northern and central regions of the country. Using a combination of morphological and molecular methods, 7 Paragonimus species, namely P. heterotremus, P. westermani, P. skrjabini, P. vietnamensis, P. proliferus, P. bangkokenis and P. harinasutai, have been identified in Vietnam. Of these, the first 3, P. heterotremus, P. westermani and P. skrjabini, are known to infect humans in other countries. However, in Vietnam, only P. heterotremus, found in some northern provinces, has been shown to infect humans. Even nowadays, local people in some northern provinces, such as Lai Chau and Yen Bai, are still suffering from P. heterotremus infection. In some provinces of central Vietnam, the prevalence and infection intensity of P. westermani metacercariae in freshwater crabs (the second intermediate hosts) are extremely high, but human cases have not been reported. Likewise, although P. skrjabini was found in Thanh Hoa Province, its pathogenicity to humans in Vietnam still remains uncertain. The results of molecular phylogenetic analyses of Vietnamese Paragonimus species provides new insights on the phylogeny and taxonomy of the genus Paragonimus. Comprehensive molecular epidemiological and geobiological studies on the genus in Vietnam and adjacent countries are needed to clarify the biodiversity and public health significance of the lung flukes.
INTRODUCTION
Paragonimiasis is a food-borne parasitic zoonosis caused by infection with lung flukes of the genus Paragonimus and is a serious public health issue in some endemic areas. Paragonimus and paragonimiasis, therefore, remain a focus of research in actual or potential endemic areas [1]. New Paragonimus species and new endemic foci of paragonimiasis are still being discovered [2-4].
In Vietnam, Paragonimus and paragonimiasis have become a public health issue in the last 2 decades [5-7]. Until 2006, only 1 species, P. heterotremus, was known to occur in Vietnam and is the only confirmed species to cause human infections [8]. Since 2006, large scale investigations for Paragonimus and paragonimiasis have been conducted in northern and central Vietnam as a collaborative research between Japanese and Vietnamese parasitologists. To date, at least 7 Paragonimus species have been discovered in the country. This series of discoveries has brought new insights on taxonomic status, genetic variation, and phylogenetic relationships of Paragonimus species in Asia, which will be briefly reviewed here.
HISTORICAL BACKGROUND
From 1906 until 1995, P. westermani, the widespread Asian lung fluke, was considered to be the only lung fluke present in Vietnam and was assumed to be the cause of human paragonimiasis. However, there was no solid evidence regarding the identity of the species responsible. No morphological investigation of metacercariae or adults had been done [9,10]. In 1994 [5], endemic paragonimiasis was found in Sin Ho district, Lai Chau Province, northern Vietnam [6]. From 1995 to 1997, 2 species, P. heterotremus and P. ohirai, were identified in this area on the basis of morphology [11,12]. However, subsequent surveys in the northern provinces, including Lai Chau, found only P. heterotremus metacercariae in mountain crabs [6,7,13]. Since then, we have been conducting extensive surveys for Paragonimus species in northern and central Vietnam using a combination of morphological and molecular analyses for metacercariae from crab hosts and adults obtained by experimental infections. So far, 7 Paragonimus species have been found in Vietnam. In order of discovery, these are P. heterotremus, P. vietnamensis, P. proliferus, P. bangkokensis, P. westermani, P. harinasutai, and P. skrjabini [2,14-19].
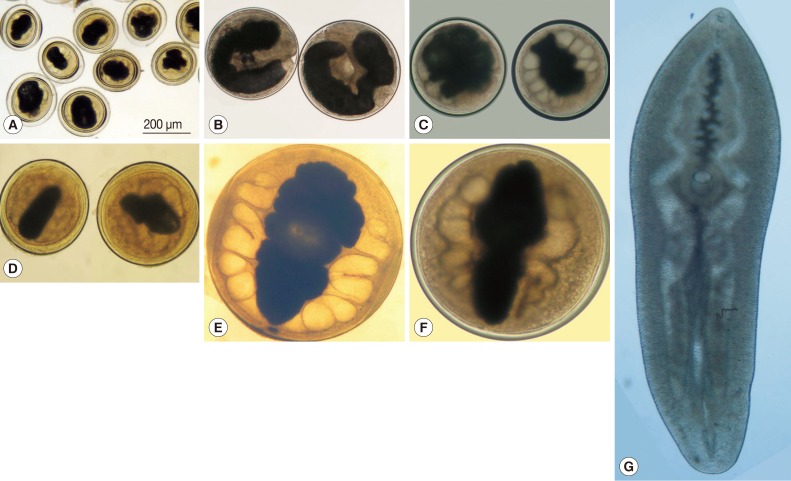
MORPHOLOGICAL FEATURES OF PARAGONIMUS SPP. IN VIETNAM
Except for the adults of P. heterotremus found in natural hosts [6], adult worms of all other Paragonimus species were collected from laboratory animals by experimental infection with metacercariae isolated from the second intermediate host crabs. Since metacercariae of more than 1 species were sometimes found in the same individual crab host in the same sampling area [14-19], the identification of metacercariae requires experience and good eyes. Here are some keys to species identification.
KEY TO IDENTIFICATION OF PARAGONIMUS METACERCARIAE IN VIETNAM
1a. Metacercariae usually found excysted (about 2.5×0.7 mm) in crabs ... P. proliferus (Fig. 1G)
b. Metacercariae usually found encysted form in crabs ... 2
2a. Metacercariae <300 µm in diameter ... P. heterotremus (Fig. 1A)
b. Metacercariae >300 µm in diameter ... 3
3a. Metacercariae about 400-500 µm in diameter ... 4
b. Metacercariae about 700-800 µm in diameter ... 6
4a. Cyst walls thin; metacercariae U-shaped in cyst, not occupying entire cyst cavity ... P. bangkokensis (Fig. 1B)
b. Cyst walls thick; metacercariae fully occupy cyst cavity ... 5
5a. Metacercariae relatively translucent ... P. westermani (Fig. 1C)
b. Metacercariae relatively turbid ... P. skrjabini (Fig. 1D)
6a. Margins of excretory bladder wrinkled ... P. vietnamensis (Fig. 1E)
b. Margins of excretory bladder relatively smooth ... P. harinasutai (Fig. 1F)
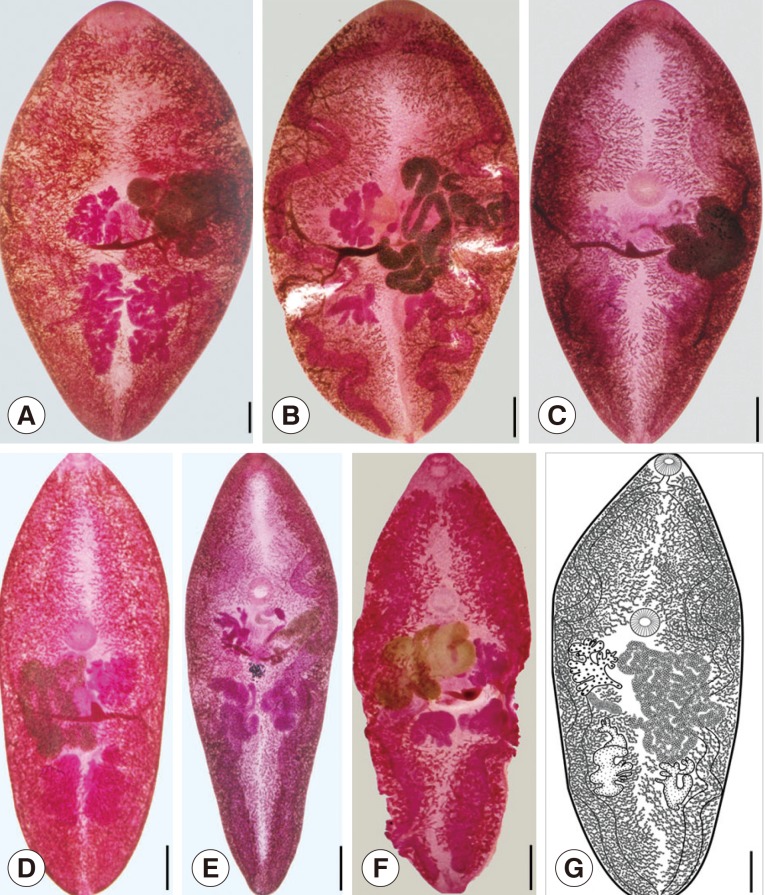
KEY TO IDENTIFICATION OF PARAGONIMUS ADULTS IN VIETNAM
1a. Body oval; body length:width ratio about 1.4-2.0 ... 2
b. Body is more elongate; body length:width ratio about 2.5-3.0 ... 4
2a. Oral sucker nearly twice diameter of ventral sucker ... P. heterotremus (Fig. 2A)
b. Oral sucker almost equal to ventral sucker in diameter ... 3
3a. Ovary and testes have 5-6 lobes ... P. westermani (Fig. 2B)
b. Ovary and testes finely branched, not lobed ... P. vietnamensis (Fig. 2C)
4a. Spines grouped on body surface ... 5
b. Spines single on body surface ... 6
5a. Posterior end of body more tappered; the ventral sucker and the reproductive organs are located more posteriorly ... P. bangkokensis (Fig. 2D)
b. Posterior end of body is relatively pointed; ventral sucker and reproductive organs located more anteriorly ... P. proliferus (Fig. 2E)
6a. Testes not extensive; occupy about 1/10 the body length ... P. harinasutai (Fig. 2G)
b. Testes extensive; occupy about 2/10 the body length ... P. skrjabini (Fig. 2F)
GEOGRAPHICAL DISTRIBUTION
Geographical distribution of the 7 Paragonimus species in Vietnam is summarized in Fig. 3. Metacercariae of P. heterotremus have been found in 8 northern provinces where human paragonimiasis cases have also been recorded [7]. In addition, metacercariae of this species have been found in Thanh Hoa Province, in northern central Vietnam, but human cases have not yet been found in this province. In contrast, P. westermani was found in 3 central provinces, but no human cases have been detected so far in that region. P. bangkokensis and P. proliferus were found in both north and central provinces. The remaining species, P. skrjabini and P. harinasutai, are rare.

Geographical distribution of Paragonimus species in northern and central Vietnam.
As for P. ohirai in Vietnam, Le et al. [12] reported adults of this species in 1997 from a pig in Sin Ho district, Lai Chau Province, a mountainous area of northern Vietnam. In that study, P. ohirai was identified based on the presence of grouped spines on the worm surface and possession of a ventral sucker that was larger than the oral sucker. However, these characters are also seen in other species, P. bangkokensis and P. proliferus, of which metacercariae were also discovered in Sin Ho district, Lai Chau Province [15,16]. Metacercariae of P. ohirai have never been found in freshwater crabs in Vietnam. Moreover, P. ohirai has been reported only in the coastal regions of Japan, Korea, China, and Taiwan [1]. It is, thus, highly likely that the specimens identified as P. ohirai by Le et al. [17] in 1997 were P. bangkokensis or P. proliferus. Unfortunately, the few specimens collected by Le et al. are not in a good condition for morphological re-examination or molecular study for the accurate re-identification. Whether P. ohirai exists in Vietnam or not needs further clarification.
INTER- AND INTRA-SPECIES VARIATION AND MOLECULAR PHYLOGENETIC RELATIONSHIP
The nuclear ribosomal second internal transcribed spacer region (ITS2) is a good marker for inter-species variation, whereas partial mitochondrial cytochrome oxidase subunit 1 gene (CO1) is a good marker for intra-species variation [20]. We have determined ITS2 and CO1 sequences of all Paragonimus species in Vietnam and these are now available in GenBank [14-19].
Since it is difficult to identify some closely related Paragonimus species based on morphology alone, molecular analyses are necessary and important, especially for the description of a new species. P. vietnamensis was described as a new species in 2007 [2]. It is morphologically somewhat similar to several others, such as P. harinasutai, P. microrchis, and P. skrjabini, in either metacercarial or adult stages. However, molecular phylogenetic analyses clearly showed it to be genetically distinct from any other known Paragonimus species [14]. To date, P. vietnamensis has been found in crabs collected in 4 provinces and appears to be genetically rather homogeneous.
Likewise, P. heterotremus populations from different localities in Vietnam showed high genetic similarities with each other and clustered with those from China and Thailand in phylogenetic trees [8,14]. Based on the ITS2 sequences of eggs collected from patients, human infection with P. heterotremus in Vietnam has been confirmed [8,21].
Molecular studies have also provided solid evidence for the presence of P. westermani in Vietnam. Attention should be paid to this species because of its wide distribution and its importance as a cause of human paragonimiasis in northeastern Asian countries and the Philippines [1]. P. westermani is genetically diverse and has been considered as a species complex including 2 groups: Northeast Asia (Japan, Korea, China, and Taiwan) and Southeast Asia (Malaysia, the Philippines, and Thailand) [22,23]. The recent discovery of P. westermani in Vietnam, India, and Sri Lanka provided evidence that much remains to be learned about the P. westermani species complex [17,24,25]. P. westermani populations found in central Vietnam have high molecular similarities with those of the northeastern Asian group rather than the southeastern Asian group. However, unlike P. westermani in northeastern Asia, Vietnamese P. westermani seems not to utilize feline or canine final hosts [17], and no human cases due to this species have been found in Vietnam [21]. P. westermani recently found in India and Sri Lanka are far distant from other populations [24,25]. Blair et al. [26] first proposed in 2001 an evolutionary scenario, whereby populations of P. westermani arose first in Southeast Asia and utilized thiarid snails. Later, by range expansion and the addition of pleurocerid snail hosts, they were able to establish populations in East Asia. More recently, Devi et al. [27] in 2012 considered that the complex arose in southern Asia, perhaps in Sri Lanka, and radiated as populations were established further north and east in Asia, utilizing different snail hosts within the Cerithioidea. However, the presence of 2 distantly related members of the complex in the same place, as in Thailand and Northeast India, is not explained by such a scenario. Moreover, molecular analyses showed that genetic distances between some members of the P. westermani group are as great as between very distinct species, indicating that the taxonomy of P. westermani complex is more complicated than previously thought. The identities of snail hosts of members of the complex across the range may help to make clear the evolution of this group of lung flukes.
Another species complex, the P. skrjabini complex, consists of 2 subspecies, P. skrjabini skrjabini and P. skrjabini miyazakii [28]. Recently, P. skrjabini was found in India [29] and Vietnam [19]. Genetically, the P. skrjabini population from Vietnam was almost completely identical with those from Yunnan, China [19]. Remarkably, these Vietnamese and Chinese Yunnan populations, together with the Indian P. skrjabini population, form a distinct clade within the P. skrjabini complex. In addition, P. proliferus should also be placed in the P. skrjabini complex [19, 28]. This is a rare species originally reported in China [30]. Recently, we found P. proliferus metacercariae in Lai Chau Province in northern Vietnam [15] and Quang Binh Province in central Vietnam [31]. By molecular phylogenetic analysis, P. proliferus in Lai Chau Province was found to have ITS2 and CO1 sequences almost completely identical with those from China, whereas P. proliferus found in Quang Binh Province was far distant (5.6% difference in CO1 sequences) from the populations of Lai Chau, Vietnam, and of China. This degree of variation suggests that P. proliferus might be widely distributed in Indochina, or even broadly in Southeast Asia. Related to this, P. hokuoensis, a species found in a limited area of China and of which metacercariae are much smaller than those of P. proliferus, was genetically completely identical with P. proliferus and was placed as a synonym of P. proliferus [15].
P. bangkokensis and P. harinasutai are morphologically distinct from each other at both adult and metacercarial stages. Metacercariae may occur in the same individual crab host [18,32-34]. Recent molecular studies provided strong evidence of their close phylogenetic relationship [16,18]. All P. harinasutai and P. bangkokensis populations from Thailand, Lao PDR, and Vietnam (and also P. microrchis from China as a synonym of P. harinasutai) are genetically highly similar to each other and appear as a species complex in both ITS2 and CO1 trees. Two types of indels (AT and ATC) were independently observed in ITS2 sequences among P. harinasutai/P. bangkokensis populations. These indels are not species-specific, but have some tendencies to associate with particular geographical populations, indicating recent gene flow between P. harinasutai and P. bangkokensis and recent divergence of these 2 species [16,18]. The recent discovery of naturally hybridized specimen between P. bangkokensis and P. harinasutai in central Vietnam [35] strongly supports their close relationship in the same species complex.
INTERMEDIATE AND FINAL HOSTS FOR PARAGONIMUS SPP. IN VIETNAM
To maintain the life cycle of Paragonimus species, at least 3 hosts are required: freshwater snails as the first intermediate host; crustaceans as the second intermediate host; and mammals as the definitive hosts. So far, there have been few reports about the first intermediate host for Paragonimus spp. in Vietnam. This may be due partly to the low prevalence of infection typical in snail hosts and to difficulties in identification of snail species. Doanh et al. [13] in 2002 found Paragonimus sp. cercariae in 2 snail species, both of which were tentatively identified as Oncomelania species, at low infection rates (0.2-0.75%). Almost at the same time, De et al. [7] in 2003 reported Melanoides sp. as the first intermediate host of P. heterotremus with the infection rate of 1.4-5.0%. A figure of the cercaria was not provided. However, experimental infection of Melanoides sp. with P. heterotremus miracidia has been unsuccessful in several studies [13,36].
All 7 Paragonimus species in Vietnam utilize mountain crabs of the family Potamidae as the second intermediate host [13-19]. Prevalences of metacercariae of P. heterotremus and P. westermani are much higher than those of the other species. The infection rate of P. heterotremus metacercariae is the highest (3.0-99.8%); followed by P. westermani (2.0-94.2%), P. proliferus (3.0-32.0%), P. bangkokensis (2.0-31.0%), P. vietnamensis (1.0-29.6%), P. skrjabini (25.0%), and P. harinasutai (4.0%). The coexistence of more than 1 Paragonimus species in the same crab host was frequently found [14-19]. These results indicated that there is no competition between Paragonimus spp. metacercariae in the second intermediate hosts.
For the definitive hosts, only P. heterotremus was found in naturally infected dogs and civets, but adult worms of all other species were obtained by experimental infection [6,7,14-19]. Dogs and cats were suitable hosts for P. heterotremus, P. bangkokensis, P. proliferus, P. vietnamensis, P. skrjabini, and P. harinasutai although the period required for maturation was variable among the species. Although dogs and cats are generally suitable hosts for P. westermani elsewhere in its range, Vietnamese P. westermani populations were hardly able to develop into mature adults in these animals. To elucidate the precise life cycle of Paragonimus species, more extensive surveys on the intermediate and final hosts of Paragonimus spp. in wildlife in Vietnam are necessary.
HUMAN PARAGONIMIASIS
The first case of paragonimiasis in Vietnam was reported in 1906 [9]. However, no much attention was paid to this disease until 1994 when a number of patients were diagnosed in Sin Ho district of Lai Chau Province, northern Vietnam [5,6]. Since then, several surveys revealed that paragonimiasis is endemic in 8 provinces of the northern mountainous areas (Fig. 3). The prevalence ranged from 0.2 to 11.3% [6,7]. The majority (66.7-86.0%) of patients were children under 15 years of age [6,7]. In spite of over 15 years of repeated mass screening, treatment, and education about paragonimiasis, Doanh et al. [21] in 2011 still found patients in some previously reported endemic areas, especially in Sin Ho district of Lai Chau Province, where seroprevalence rate was 12.7% (28/220) by multiple dot-ELISA screening test. Continuous efforts on health education are, therefore, required to encourage the local people to avoid Paragonimus infection.
CONCLUSION AND PERSPECTIVES
Paragonimus species in Vietnam are more diverse than previously thought. Using a combination of morphology and molecular studies, at least 7 Paragonimus species have been found in northern and central Vietnam. All these species use mountain crabs of the family Potamidae as the second intermediate host. Metacercariae of P. heterotremus are the commonest in northern provinces, but this species has not been found in central Vietnam. In contrast, P. westermani is dominant in central Vietnam where P. heterotremus has not yet been found. Such differences in distribution may depend on the distribution of specific first intermediate hosts, a topic which has not been studied much in Vietnam.
The results of molecular phylogenetic analyses have provided a comprehensive taxonomy of Vietnamese Paragonimus and have given new insights on the phylogeny and taxonomy of the genus. The phylogenetic structure of the genus is far more complicated than previously thought, drastically influencing our understanding of the taxonomy of Paragonimus species.
To date, only P. heterotremus is confirmed to infect humans in Vietnam, and the ability of P. westermani and P. skrjabini to infect humans in Vietnam is still questionable. Although crab hosts in central provinces are found to be heavily infected with P. westermani metacercariae, the final hosts of this species in Vietnam are unknown.
Conclusively, recent discoveries of Paragonimus spp. in Vietnam have provided important data related to the epidemiology, taxonomy, and phylogeny of the genus Paragonimus. These also raised further questions such as: Are there more Paragonimus species and endemic areas in Vietnam, especially in southern Vietnam? Do any Paragonimus spp. other than P. heterotremus infect humans? How are the life cycles of Paragonimus spp. maintained in Vietnam? Much remains to be done.