Prevalence and molecular analysis of glucose-6-phosphate dehydrogenase deficiency in Chin State, Myanmar
Article information

Abstract
Glucose-6-phosphate dehydrogenase (G6PD) deficiency is caused by X-linked recessive disorderliness. It induces severe anemia when a patient with G6PD deficiency is exposed to oxidative stress that occurs with administration of an antimalarial drug, primaquine. The distribution of G6PD deficiency remains unknown while primaquine has been used for malaria treatment in Myanmar. This study aimed to investigate the prevalence of G6PD deficiency and its variants in Chin State, Myanmar. Among 322 participants, 18 (11 males and 7 females) demonstrated a G6PD deficiency. Orissa variant was dominant in the molecular analysis. This would be related to neighboring Indian and Bangladeshi population, in which Orissa variant was also reported as the main mutation type. The screening test for G6PD deficiency before primaquine treatment appears to be important in Myanmar.
Introduction
Glucose-6-phosphate dehydrogenase (G6PD) deficiency is a highly prevalent X-linked recessive genetic disorder that affects approximately 400 million people globally. The majority of G6PD-deficient individuals appear normal. However, oxidative stressor exposure, such as foods (fava bean family and legumes), drugs (antimalarial drugs such as primaquine and sulfa drugs), and infection with microorganisms, can cause life-threatening anemia in patients with severe G6PD deficiency [1,2]. Conversely, G6PD deficiency protects against malaria by reducing parasite penetration into red blood cells (RBCs) and parasite growth in RBCs [3]. The G6PD deficiency distribution is associated with historically malaria-endemic regions such as Africa, Southeast Asia, Pacific islands (central and southern part), Southern Europe, and the Eastern Mediterranean [4–7].
G6PD catalyzes the primary reaction in the pentose phosphate pathway by oxidizing glucose-6-phosphate to 6-phosphogluconolactone through NADP+ reduction to NADPH [8]. NADPH regulates the amount of reduced glutathione by converting oxidized glutathione, and it also protects the body against free radicals. This process is important because G6PD is the only source of NADPH in RBCs [9].
The G6PD gene is located at the X chromosome’s (Xq28) telomeric region and contains 13 exons, 12 introns, and a promoter enriched in GC nucleotides. The length of the G6PD gene is 18 kb; it encodes 515 amino acids that are concealed by G6PD. The biochemical and physicochemical analysis identified >400 G6PD variants, and nearly 186 DNA sequence mutations revealed a diverse degree of G6PD deficiency [10,11]. The majority of these mutations reduced the enzyme activity level.
Myanmar is a malaria-endemic region within Southeast Asia, and primaquine is widely used to treat Plasmodium vivax and P. ovale infection and to block the gametocyte stage of P. falciparum [12]. Previous studies in Myanmar revealed that the Mahidol variant (G487A) has the highest percentage of mutations, followed by Viangchan (G871A), Canton (G1376T), Kaiping (G1388A), Chinese-4 (G392T), Union (C1360T), Mediterranean (C563T), Coimbra (C592T), Kerala–Kalyan (G949A), and Valladolid (C406T) variants, among G6PD deficiency mutations [13–16]. However, a hilly, forested, and malaria-endemic region, where data is lacking regarding G6PD deficiency, remains. Therefore, awareness of G6PD deficiency pathogenesis and distribution should be emphasized. G6PD deficiency is a genetic disorder, thus patients with deficiency must avoid oxidative stress exposure because treatment is not yet available. Thus, we investigated the prevalence of G6PD deficiency and G6PD gene variants in Chin State, Myanmar.
Materials and Methods
Ethics statement
Ethical approval was obtained from the Research Ethics Committee of Kyungpook National University (2019-0042) and the Institution Review Board, Department of Medical Research, Ministry of Health and Sports, Myanmar (Ethics/DMR/2019/131) in concordance with the Declaration of Helsinki, Council for International Organizations of Medical Sciences Guidelines, and International Conference on Harmonization in Good Clinical Practice guidelines. Informed written consent forms were obtained from participants before blood sample collection. Furthermore, parents and legal guardians of all children provided written informed consent for the use and future use of their children’s samples before enrollment into the study.
Sample collection
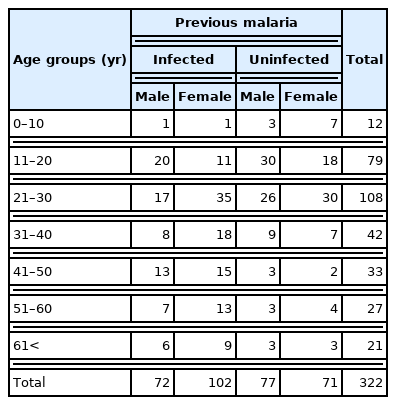
This study collected 322 blood samples from randomly selected individuals residing in Hakha City, which is the capital city of Chin State (Fig. 1). Their ethnic origin was Chin. The collection site was a mountainous region with limited transportation routes and development compared to the rest of the country. The Chin State has been consistently reported as a region with high malaria incidence since 2015 when malaria elimination programs began in Myanmar. In 2018, 64.0% of all malaria cases in Myanmar occurred in Chin State with an annual parasite incidence of 15.8 per 1,000 population [17]. Participants were aged 3–72 years, of whom 33.5% (108/322) were in the 21–30 years age group (Table 1). One or two blood spots were collected on Whatman 90-mm filter papers, allowed to air dry, and kept at −20°C for DNA extraction.
Map of blood collection regions in Myanmar (Hakha City in Chin State). Bhu.: Bhutan; Bang.: Bangladesh.
Demographic characteristics of participants in this study
Screening of G6PD deficiency
We screened for G6PD deficiency using commercially available kits according to the manufacturer’s instruction (CareStart, AccessBio, Somerset, NJ, USA). The result can be read within 20 min with the purple color shown in the reading window for G6PD deficiency. The blood was drawn with a pipette and dropped on a sample well after piercing a fingertip with a lancet. Two drops of Assay Buffer (100 μl) were added next to the buffer well immediately. The result will be available within 20 min with a noticeable purple color in the reading window for a typical result and no color change or a very dull purple color for a negative result.
Questionnaires
A questionnaire, including basic demographic data (age, gender, and ethnics), medical history (history of malaria infection and adverse malaria drug reactions), and travel history, was used to obtain information on the awareness and risk factors of G6PD deficiency.
DNA extraction
The DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany), with modifications to the manufacturer’s protocol, was used to extract genomic DNA from blood spots on filter paper. Briefly, the blood spots were dissected into a 1.5-ml microtube, and then 200 μl of phosphate buffer-saline (PBS) was added before being left overnight. The following day, 20 μl of Proteinase K, 200 μl of PBS, and 40 μl of RNase A were added, and then samples were left for 5 min at room temperature. Then, 200 μl of lysis buffer was mixed thoroughly by vortexing, before being spun down and left to rest for 2 min. After incubation at 56°C and 95°C for 15 min, 200 μl of ethanol was added, and the samples were washed with washing buffer. Finally, the samples were eluted with 50 μl of distilled water and kept at −20°C until required.
Polymerase chain reaction (PCR) and sequencing
PCR was performed in a 50-μl total volume, including 4 μl of dNTP (800 μl, 2.5 mM), 5 μl of 10×buffer, 1 μl of forward primer (10 pmol), 1 μl of reverse primer (10 pmol), 0.5 μl of exTaq polymerase (1 U/μl, TaKaRa, Tokyo, Japan), and 33.5 μl of distilled water. Supplementary Table S1 describes the primers used to amplify G6PD [18]. PCR amplification conditions were as follows: denaturation at 94°C for 5 min, followed by 40 cycles of heating at 94°C for 30 sec, the annealing temperature of 55–65°C for 30 sec, 72°C for 1 min, and a final elongation step at 72°C for 10 min. PCR products at 5 μl of were loaded using 1% and 2% gel and run for 15 min with 1 kb DNA marker (SizerTM DNA Markers, iNtRON Biotechnology, Daejeon, Korea). An AccuPrep Gel Purification Kit (Bioneer, Daejeon, Korea) was used to purify each amplicon from the gel. It was cloned into a pGEM-T TA cloning vector (Promega, Madison, WI, USA), and sequenced for each of the five colonies per the gene to avoid mis-amplification using the pUC/M13 forward and reverse primers. A Big-Dye Terminator version 3.1 Sequencing Kit (Applied Biosystems, Waltham, MA, USA) and an automatic ABI3730 DNA Analyzer Sequencer (Applied Biosystems) were used. A G6PD sequence in GenBank (accession No.X55448) was aligned as a reference sequence with the resultant nucleotide sequences to find mutations using Clustal Omega (version 2) [18]. Supplementary Table S1 describes the G6PD variants determined in this study.
Results
The CareStart G6PD deficiency test revealed G6PD deficiency in 5.6% (18/322) of the 322 screened individuals. G6PD deficiency was found in 11 males (7.4%, 11/149) and 7 females (4.0%, 7/173) (Table 2). A history of malaria was found in 6 of the 18 individuals with G6PD deficiency. Moreover, according to age, 6 (7.6%, 6/79), 7 (6.5%, 7/108), and 5 (11.9%, 5/42) individuals had G6PD deficiency in the age groups of 11–20, 21–30, and 31–40 years, respectively.

Prevalence of G6PD deficiency in Chin State, Myanmar
All 322 samples were used for amplification with primer sets of EX3-4F/R, EX5F/R, EX6-7F/R, EX9F/R, and AF3-AR3 in the PCR. Sequencing results revealed 44 G6PD variants from 40 individuals of Chin ethnicity, including 4 participants with double mutations. The variants appeared in 20 females (11.6%, 20/173) and 20 males (13.4%, 20/149). Two males and two females had double mutations in the G6PD gene. Among the 18 G6PD deficient individuals, 12 (66.7%, 12/18) had 14 mutations, whereas the remaining mutations were found in 28 individuals without G6PD deficiency. Moreover, no other novel mutations were detected in the other 6 out of 18 individuals with G6PD deficiency. The G6PD variants were found in 6 males and 6 females, respectively. Furthermore, double mutations were observed in 2 female participants with G6PD deficiency. The Orissa mutation type (n=21) was prevalent among the Chin population, followed by Chinese-4 (n=6), Mahidol (n=5), Kalyan–Kerala (n=4), Mediterranean (n=3), Viangchan (n=3), and Coimbra (n=2) mutation types (Table 3). Double mutations were as follows: Orissa and Chinses-4, Mahidol and Viangchan, Kalyan–Kerala and Viangchan, and Mediterranean and Coimbra mutation types. The G6PD deficiency phenotype revealed 14 (31.8%) and 30 (68.2%) mutation types in 12 G6PD deficient and 28 healthy individuals, respectively. Orissa’s mutation type was most predominant in both groups, G6PD deficient (n=6) and healthy (n=15) groups. However, the Mahidol variant was more frequent in G6PD deficient (n=3) than in healthy (n=2) individuals. In contrast, Chinese-4, Mediterranean, and Viangchan variants were prevalent among healthy individuals.
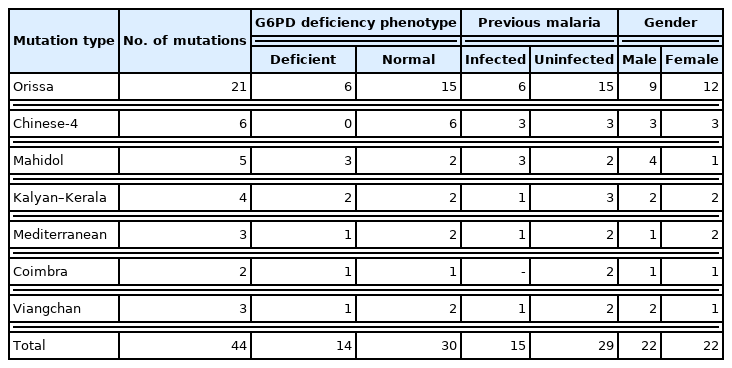
G6PD mutation type according to G6PD deficiency and malaria history
The variants were analyzed against responses recorded during the questionnaire administration to determine the association between G6PD variant distribution and malaria (Table 3). Among the participants, 174 individuals had been diagnosed with malaria previously using malaria-specific rapid diagnostic test kits (Table 1). The group with a history of malaria infection demonstrated 15 variants, whereas the group without a malaria history demonstrated 29 variants. In detail, Orissa type (6 and 15 individuals with and without malaria history, respectively) was dominant, followed by Chinese-4 (n=3) and Mahidol (n=3) mutation types in the group with malaria history and Chinese-4 (n=3) and Kalyan–Kerala (n=3) types in the group without any history of malaria.
The highest percentage of individuals with G6PD mutations (19.0%, 4/21 cases) was found in the age group of >60 years, followed by 0–10 years (16.7%, 2/12 ). Additionally, 13.9% (11/79), 13.9% (15/108), 9.5% (4/42), 6.1% (2/33), and 7.4% (2/27) of individuals with G6PD variants were seen in the age groups of 11–20, 21–30, 31–40, 41–50, and 51–60 years, respectively.
Discussion
This study revealed a previous medical history of malaria infection, either singly with P. falciparum or P. vivax or as a mixed infection with the 2 species in 174 of the 322 participants (72 males and 102 females), proving that the Chin State remains a malaria-endemic area. However, information regarding the drug administered and the participants not tested for G6PD deficiency is unavailable. The G6PD detection kit revealed G6PD deficiency in 5.6% (n=18) of the tested group. G6PD deficiency was widespread throughout the population of the Greater Mekong Subregion, including Myanmar. Previous studies determined that people from Myanmar have the second highest percentage of G6PD mutation (15.8%), with people from Cambodia having the highest percentage (18.8%), among neighboring countries, including Thailand, Lao PDR, and Vietnam [19]. The overall prevalence of G6PD deficiency (5.6%, 18/322) was lower in Chin State than in previous studies. Previous publications revealed the overall percentage of G6PD deficiency as 17.5% in Shan State, 6.6% in the Yangon Division, 13.7% in the Thai-Myanmar border area, 29.6% in China–Myanmar border area, and 11% in a remote Myanmar area [15,20,21]. Furthermore, previous studies revealed extremely diverse G6PD deficiency distribution differences according to gender. The China–Myanmar border revealed a similar distribution of 27.9% vs. 30.6% in males vs. females, whereas the G6PD deficiency distribution was relatively high among males in Yangon or Kachin State (males vs. females: 11.7% vs. 2.2% in Yangon and 9.4% vs. 2.1% in Kachin State) [15,22,23]. In contrast, a study conducted in Mandalay State reported cases with a higher G6PD deficiency distribution among females (male vs. female=4.2% vs. 8.1%) [16]. The present study revealed higher G6PD deficiency prevalence among males (7.4%, 11/149) than females (4.0%, 7/173).
Populations within Southeast Asia demonstrate a relatively high molecular heterogeneity of G6PD and various mutation distribution were found within geographical areas and/or ethnic groups [24]. G6PD mutation pattern investigated in Kinh ethnics in Vietnam revealed similarities to the Thailand population, except for the absence of Mahidol mutation while Mahidol was dominant in Southeast Asia [25]. Moreover, Viangchan, Mahidol, Mediterranean, and Canton G6PD mutations were highly prevalent in Southeast Asia, although they were very rare or completely absent in Eastern Indonesia populations [26]. Chin State has a small population, ranking among the lowest three in Myanmar, and resided by many sub-ethnic groups in Chin State [27]. This study collected samples from the Chin ethnic group, and Orissa variants were the most common. The Mahidol variant has been primarily reported in other ethnic groups of Myanmar to date, making this an interesting finding [16,22,23,28]. It can be correlated with a neighborhood of Chin State and India as well as Bangladesh. Common and widespread G6PD mutation types in India include the Orissa, Kalyan–Kerala, and Mediterranean variants [29,30]. Moreover, Orissa, Mahidol, and Kalyan–Kerala G6PD variants were mainly reported in the Bangladeshi population [31]. The Mediterranean mutation type was reported to cause a total loss of enzyme activity and neonatal jaundice, but the Orissa type is a critical mutation associated with NADPH binding site and an increased rate of neonatal jaundice caused by four residues above G6PD Aures [32]. The residues involved in binding NADP in G6PD are a variant of the dinucleotide-binding fingerprint GXGXXG or GXGXXA, namely GASGDLA, between residues 38 and 44. The Orissa G6PD variant has a glycine instead of an alanine residue at position 44. A previous report revealed that the alternative residue, glycine, results in significantly decreased coenzyme binding and decreased efficiency of G6PD Orissa, inducing RBC property alterations that are resistant to malaria infection [30,33,34].
The present study mainly revealed a single G6PD mutation type per individual, but four double mutation types were also reported. This observation could raise concerns regarding occurrence of severe G6PD deficiency, because double mutations, such as Viangchan and Mahidol double mutant, induced nearly 10-fold less enzyme activity than a single mutant and were illustrated as a severe enzyme deficiency in a previous study [35].
To date, studies on G6PD deficiency prevalence and G6PD variants have been conducted on several ethnic groups in Myanmar, including Burmese, Kachin, Rakhine, Shan, and Chin [13–16,20–23,28]. The highest G6PD deficiency prevalence was found in a study of the Kachin population, wherein the G6PD deficiency distribution was 30.5% [15]. Another study on individuals of Chin ethnicity revealed a 9% G6PD deficiency distribution [28]. Moreover, studies of G6PD genotyping revealed that the Mahidol variant was dominant in Myanmar [13–15,20–23]. The present study G6PD enzyme deficiency in 5.6% of the Chin population. These data are lower than the mean rate in other regions of Myanmar. The molecular analysis revealed that the Orissa mutation type was dominant, followed by Chinese-4, Mahidol, Kalyan-Karala, Mediterranean, Coimbra, and Viangchan. This result is different from the previous results revealing that the Mahidol type was primarily detected in Myanmar. However, these data would be associated with neighboring Indian and Bangladeshi populations having the Orissa type as the main mutation. The information from this study is important, considering that the public health concern of G6PD deficiency remains very weak, and malaria incidence remains on the rise. The screening test before taking antimalarial drugs and newborn screening for G6PD deficiency is strongly recommended in this area, and a follow-up investigation of the toxicity of antimalarial drugs would be required.
Supplementary Information
Acknowledgments
We acknowledge all the blood sample givers and all the staff from Chin Christian University who helped throughout the survey process. This research was financially supported by the Basic Science Research Program (NRF-2019R1C1C1002170) and the framework of the international cooperation program (2022K2A9A1A01098057, FY2022) funded by the Ministry of Science, ICT and Future Planning through the National Research Foundation of Korea (NRF).
Notes
The authors declare no conflict of interest related to this study.
Author contributions
Conceptualization: Hong Y, Chung DI, Goo YK
Data curation: Aung JM, Moon Z, Dinzouna-Boutamba SD
Formal analysis: Aung JM, Moon Z
Funding acquisition: Goo YK
Investigation: Aung JM, Moon Z, VanBik D
Resources: Aung JM, VanBik D, Ring Z
Validation: Lee S, Goo YK
Visualization: Lee S
Writing-original draft: Aung JM, Goo YK
Writing-review& editing: Aung JM, Lee S, Goo YK