Introduction
Trichinella spiralis is a zoonotic nematode that causes trichinellosis (or trichinosis) in humans and can infect various mammals worldwide [1]. The parasite can be transmitted by consuming raw or undercooked infected meat, especially pork muscle that contains larvae (TsL1). After exposure to gastric juice in the stomach, TsL1 larvae are released from the muscle tissue and then penetrate the small intestinal wall and develop into adult worms. Adult males and females mate, after which the females release newborn larvae (NBL) that migrate via the blood or lymphatic circulation to muscles and particularly to highly oxygenated skeletal muscles [2]. To survive long-term in their hosts, T. spiralis larvae establish chronic infections in their host muscles by modulating and escaping potentially destructive immune responses. This is achieved by anatomical seclusion involving intracellular infection and nurse cell formation, as well as regulating host immune responses by releasing immunomodulatory molecules found in excretory–secretory products (ES) [2,3]. In some instances, TsL1-ES can modulate the immune response primarily through the TLR4-dependent pathway by decreasing the expression of this receptor [4]. The incubation of TsL1-ES with HEK cells transfected with mouse TLR4 (TLR4/MD2-CD14) can suppress LPS-mediated TLR4 activation, reducing proinflammatory cytokine production [5].
In our previous study, recombinant T. spiralis novel cystatin (rTsCstN) inhibited the activity of cathepsin L (CatL) in vitro and ex vivo in mouse bone marrow-derived macrophages (mBMDMs). Moreover, rTsCstN exhibited anti-inflammatory properties in mBMDMs by suppressing lipopolysaccharide (LPS)-mediated proinflammatory cytokines, including tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and interferon (IFN)-γ and by interfering with the antigen presentation process through the suppression of MHC class II expression [6]. In addition, the interaction between TsCstN-mBMDMs impaired the Th1 response, affecting downstream T-cell priming by lowering IFN-α and IL-12 levels and interfering with STAT4 signaling [7]. To achieve an in-depth understanding of properties and functions of TsCstN, the protease inhibitory domain of the protein should be identified and characterized and establish if future modifications or truncation of this protein could increase its activity and specificity against inflammatory diseases.
In previous studies, cystatin-derived peptides have been designed and used for therapeutic purposes, including anti-inflammatory, antimicrobial, and anticancer effects. The administration of peptides derived from filarial cystatin to mice with dextran sulfate sodium–induced colitis significantly reduced gross and histological pathological alterations and inflammatory responses [8]. The N-terminal fragment of human cystatin C exhibited strong antibacterial action against several bacterial species, including methicillin-resistant S. aureus, frequently isolated from infected wounds [9]. A small cystatin C peptide also demonstrated anticancer effects by decreasing cell growth, proliferation, and migration and increasing apoptosis in melanoma cells in vitro [10].
In this study, we identified the protease inhibitory reactive regions of TsCstN by using an in silico truncation modeling approach. Full-length TsCstN was employed to simulate secondary (2D) and tertiary (3D) structures before determining the truncation locations based on the functional domains of the proteins. Subsequently, recombinant proteins of TsCstN and its truncates were expressed in a prokaryotic expression system or artificially synthesized, followed by an evaluation of their properties through the inhibition of human CatL. The results of this study will help in the future development of biotherapeutic medications for immune-mediated disorders and chronic inflammatory diseases.
Materials and Methods
Bioinformatic approach to design TsCstN protein truncates
In our previous study, TsCstN was shown to lack conserved cystatin sequences according to multiple protein alignments with its homologs [6]. Therefore, we gathered additional cystatins derived from various helminths to reassess the multiple alignment and validate our earlier discovery [6]. Amino acid sequences of ten helminth cystatins were retrieved from the NCBI database (https://www.ncbi.nlm.nih.gov/genbank/); amino acid sequences and accession numbers are provided in Supplementary Table S1. Multiple sequence alignment was performed to compare the conserved sequence regions using ClustalX2 [11] and visualized using the Bio-Edit alignment editor [12].
Design of TsCstN truncates and protein structure simulation
The amino acid sequence of full-length TsCstN (Ts01) was used to determine the protein 2D structure predicted by five different programs: Jpred4 [13], PHD [14], Phyre2 [15], PSIPRED [12], and PredictProtein (https://predictprotein.org/). The TsCstN truncates, TsCstNΔ1–39 (Ts02), TsCstNΔ1–71 (Ts03), TsCstNΔ1–20, Δ73–117 (Ts04), and TsCstNΔ1–20, Δ42–117 (Ts05), were manually designed by removing protein domains based on the 2D structures and functional protein domains of Ts01 to generate Ts02–Ts05 were subsequently predicted as 2D structures using the same programs as above for Ts01 to confirm structure change and stability after truncation; predicted structures were then aligned using the Bio-Edit alignment editor [16]. The molecular weight and isoelectric point of Ts01–Ts05 were predicted using the ProtParam tool on the ExPASy server (http://www.expasy.org/tools/).
I-TASSER [17] and the AlphaFold2 method [18] were used to simulate the 3D structure of Ts01–Ts05. The surface area, volume, and 3D structures of these proteins were visualized using Biovia discovery studio (Discovery Studio Modeling Environment, Release 2022. Dassault Systèmes; San Diego, CA, USA: 2021), and the Ramachandran plot was evaluated to verify the quality of each structure using PROCHECK analysis [19]. Structures of truncated TsCstN were superimposed onto Ts01 using the PyMOL2 server (http://www.pymol.org/pymol). The overall conceptual framework of protein modeling is shown as a flow chart (Supplementary Fig. S1).
DNA amplification and cloning of truncated TsCstN DNAs
pET23b+ carrying full-length TsCsN without a signal peptide (pET23b-Ts01) was prepared in our previous publication [6] and was used as the DNA template for amplifying truncated TsCstN (Ts02–Ts04) DNAs. The specific primers were designed to amplify the encoding DNA region of each truncate, and the BamHI and HindIII restriction sites were incorporated into the 5ºC-end of the forward (fw) and reverse (rv) primers, respectively, for cloning into pET23b+. Specific primers for amplifying Ts01–Ts04 are listed in Supplementary Table S2. The Ts05 peptide was synthesized by GenScript (Piscataway, NJ, USA).
The truncated TsCstN was amplified via conventional PCR in a total volume of 20 μl containing 2 μl of pET23b-Ts01 plasmid (approximately 1 ng), 1× Dream Taq DNA polymerase buffer, 0.2 mm of each dNTP, 1 U of Dream Taq DNA polymerases, and 100 nM of Fw- and Rv-specific primers of each truncate. PCR conditions consisted of 94ºC for 5 min, 35 cycles of 94ºC for 30 sec, 55ºC for 30 sec, and 72ºC for 2 min and a final extension step at 72ºC for 5 min. Results were checked via 3% gel electrophoresis. The target amplicon was excised from the gel and purified using a GenepHlow Gel/PCR kit (Geneaid Biotech, New Taipei, Taiwan). The pET23b+ plasmid (Novagen-EMD Biosciences, Darmstadt, Germany) and purified Ts02, Ts03, or Ts04 amplicons were digested with BamHI and HindIII at 37ºC for 4 h before ligation with T4 DNA ligase (Thermo Fisher Scientific, Waltham, MA, USA). The ligation reaction was transformed into Escherichia coli strain JM109 via heat-shock. The following day, 5 to 10 single colonies from each plate of truncated TsCstN were selected to confirm and validate DNA recombination using colony PCR, restriction endonuclease digestion, and DNA sequencing.
Expression and purification of full-length and truncated TsCstN proteins
Recombinant truncated TsCstN, the plasmids pET23b+ carrying Ts02, Ts03, or Ts04 (pET 23b-Ts02, pET23b-Ts03, and pET23b-Ts04, respectively) were isolated from E. coli strain JM109, and transformed E. coli strain BL21 (DE3). BL21 cells carrying plasmid pET23b-Ts01, -Ts02, -Tso3, or -Ts04 were used to express and purify recombinant proteins as previously described [6]. Recombinant mouse dihydrofolate reductase (rmDHFR) was produced using the procedures outlined in our previous study and employed as an irrelevant protein control [6].
Protease inhibitory assay
The Ts01–Ts05 proteins were evaluated for their inhibition of human CatL activity (Sigma-Aldrich, St Louis, MO, USA). rTs01 (129 μM), rTs02 (392 μM), rTs03 (243 μM), rTs04 (350 μM), and rTs05 (19.6 mM) were diluted to 10, 20, or 40 μm and incubated with assay buffer (100 mM sodium acetate, pH 5.5, 100 mM NaCl, 1 mM DTT) at 37°C for 30 min. Subsequently, the reaction was added to Nunc F96 MicroWell Black plate, followed by the addition of 0.5 nM CatL into the well and incubation at 37°C for 30 min. Then, 100 μl of ta fluorogenic CatL substrate (10 mM of Z-Phe-Arg-AMC-HCl) (Chem-Impex International, Wood Dale, IL, USA) was added per reaction. The fluorescence intensity was monitored at 37°C for 30 min using a fluorometer (Tecan Infinite M Plex, Tecan Austria GmbH Untersbergstr. 1A, A-5082 Grödig, Austria) with excitation and emission at 360 and 480 nm, respectively. The percentage of residual enzyme activity was determined. rmDHFR and E64 were used as irrelevant and positive inhibitor controls, respectively. Experiments were performed in duplicate with two independent experiments.
Statistical analysis
Statistical analyses were performed using GraphPad Prism 7 software (GraphPad, La Jolla, CA, USA). The significance between each group was analyzed using one-way ANOVA followed by multiple comparison tests. Individual data in each group are presented as the mean±SD; a P-value <0.05 was considered significant.
Results
Designing TsCstN truncates using a nonhomology structure strategy
A multiple sequence alignment of TsCstN (Ts01) with related cystatin homologs was performed using ClustalX2. The Ts01 amino acid sequence had a low conservation level. Compared with cystatin homologs, Ts01 did not contain any of the cystatin signature motif present in cystatin homologs (Supplementary Fig. S2). The structure template for the 3D protein simulation of Ts01 was obtained from the SWISS-MODEL server (https://swissmodel.expasy.org/interactive PDB) [20], although the PDB database currently lacks a structural homolog for Ts01. Therefore, it was not possible to simulate Ts01 or create protein truncates using comparative homology modeling.
To design the truncate TsCstN proteins, the Ts01 2D structures were predicted with pred4, PHD, Phyre2, and PSIPRED. The Ts01 2D structure and functional protein domain were used to predict the sites for truncation locations that would produce TsCstN truncates. Four distinct protein truncates, Ts02, Ts03, Ts04, and Ts05, were generated in this study. Following a comparison with the Ts01 structure, the 2D structures of the truncates were predicted in the same manner as for Ts01. The 2D structures of the truncates aligned with Ts01 from all from five different algorithms (Supplementary Fig. S3). The amino acid sequences, 2D structure composition of Ts01, truncates, molecular weights (kDa), and isoelectric point are summarized in Supplementary Table S3.
3D structural simulation of TsCstN truncates to evaluate conformational alteration
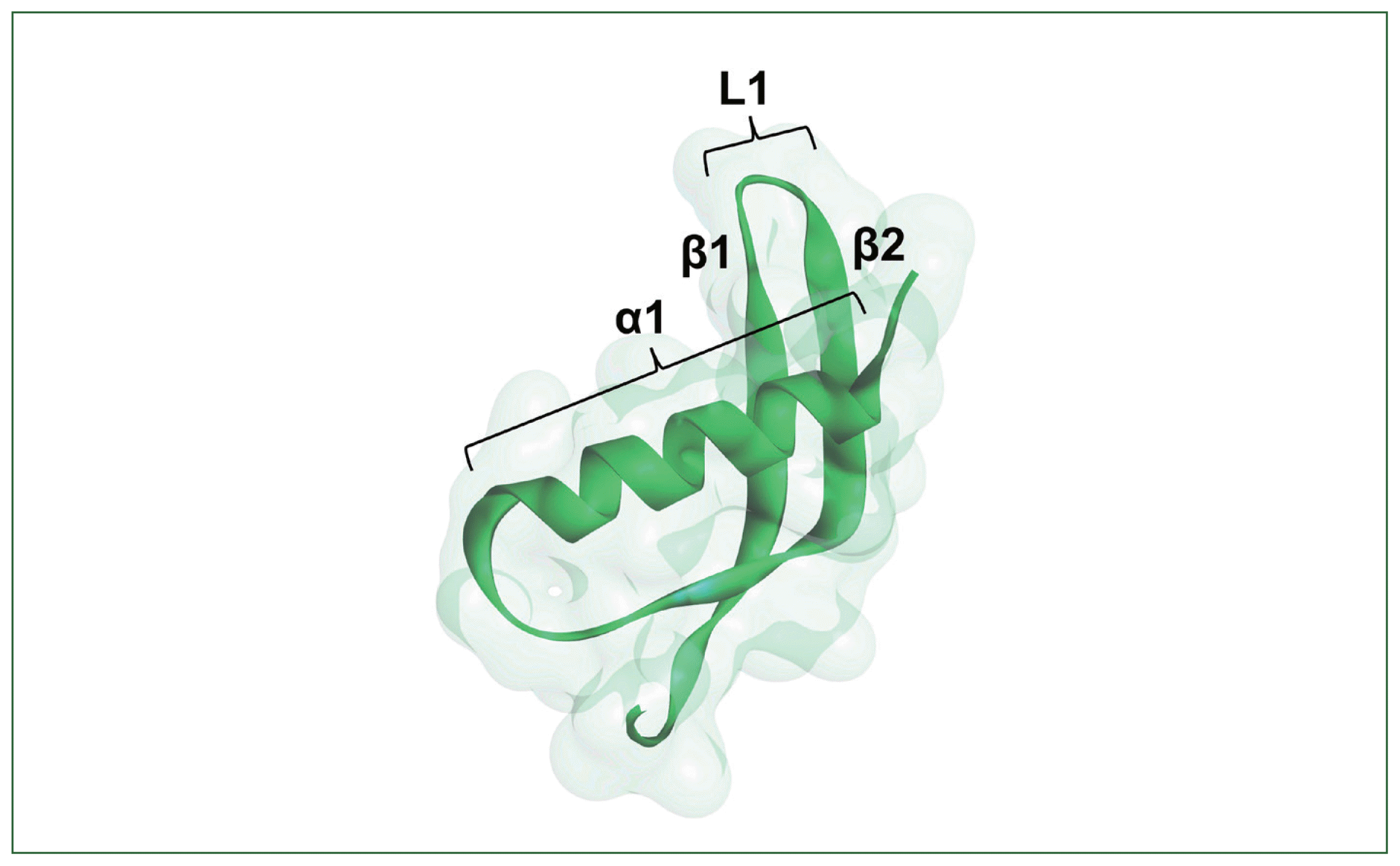
I-TASSER and Alpha-Fold2 were used to model Ts01 3D structures and truncates, with both techniques predicting all structures comparably. In detail, Ts01 contained a full α1 helix (α1), loop1 (L1; β1β2), α2 helix (α2), an irregular coil, and Loop2 (L2; β3β4) (1–117 aa); Ts02 contained L1, α2, an irregular coil, and L2 without α1 (40–117 aa); Ts03 contained α2, an irregular coil, and L2 without α1 or L1 (72–117 aa); Ts04 contained α1 and L1 (21–72 aa); and Ts05 refers to α1 structure only (21–41 aa) (Supplementary Figs. S4, S5). Results from the Ramachandran plots and the supporting information of protein structure modeling, such as surface area and volume, were also used for analysis. The structures of proteins showing a favored region pf >70% and a disallowed region pf <2% were selected to align against Ts01 to select the closely related structure that was similar to the predicted structure. Based on the Ramachandran plot, the findings indicated that I-TASSER exhibited a lower precision in predicting 3D protein structure. Most protein structures displayed a small percentage of favored regions (<70%) and a significantly higher percentage of disallowed regions (>2%). Conversely, AlphaFold2 demonstrated a higher accuracy in predicting 3D structures, featuring a notable percentage of the favored regions (>70%) and a minimal percentage of the disallowed regions (<2%). The 3D structure modeling of Ts04 demonstrated the best quality with the highest percentage of favored regions (Supplementary Tables S4, S5).
To validate the results of the 3D structure simulation, the truncated TsCstN (Ts02–Ts05) structures were superimposed on Ts01. The proteins modeled by AlphaFold Colab had a structure more comparable with that of Ts01 than those modeled with I-TASSER (Supplementary Figs. S6, S7). The structure of TS04 was the most comparable with that of Ts01 when modeled with AlphaFold2 (Supplementary Fig. S7C).
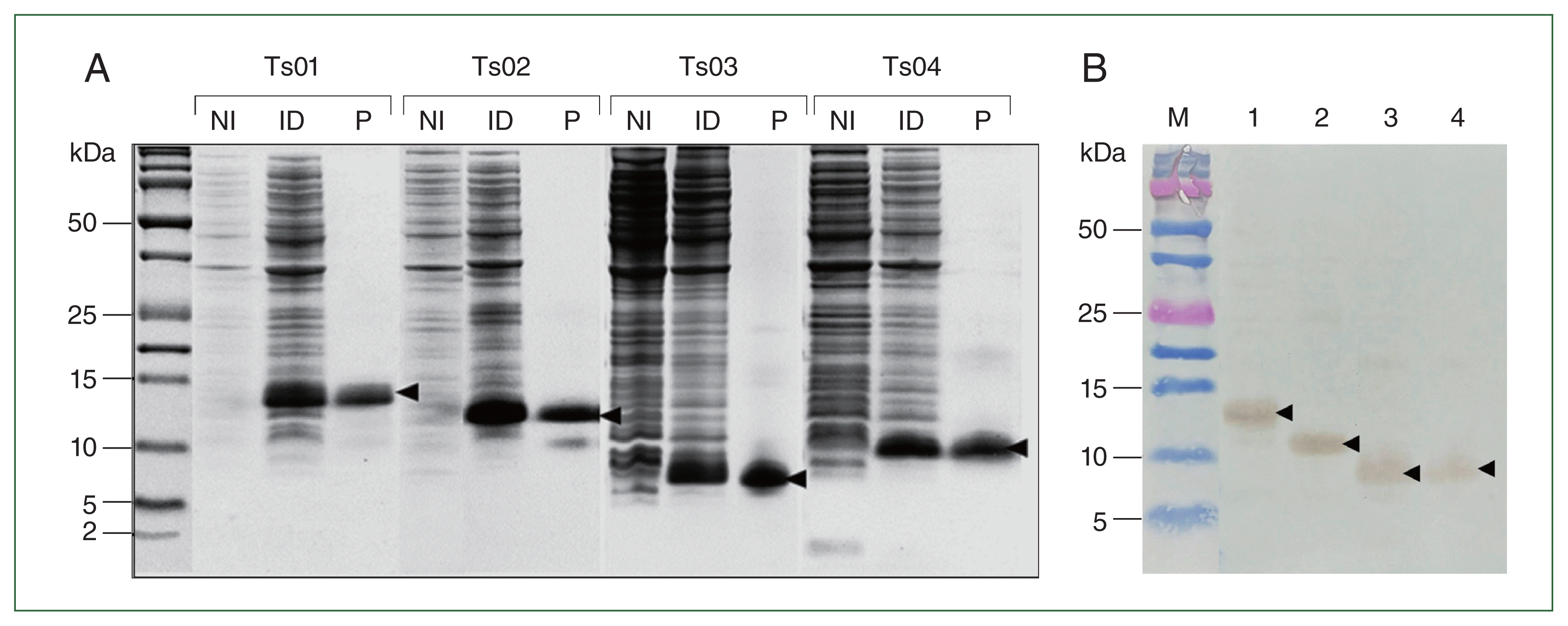
Production of recombinant proteins for functional analysis
Recombinant Ts01, -Ts02, -Ts03, and -Ts04 were overexpressed in E. coli in an insoluble form at molecular weights of approximately 14, 12, 8, and 10 kDa, respectively (Fig. 1A). Purified recombinant proteins were completely refolded and solubilized in 1× PBS before the protein concentrations were measured. Western blot analysis of rTs01, rTs02, rTs03, and rTs04 with mouse anti-His Tag antibodies confirmed that the antibody reacted with all recombinant proteins (Fig. 1B). rmDHFR was used as an irrelevant control protein and was expressed in a soluble form at a molecular weights of approximately 24 kDa and subsequently purified under native conditions (Supplementary Fig. S8).
Evaluation of the protease inhibitory activity of TsCstN and truncates
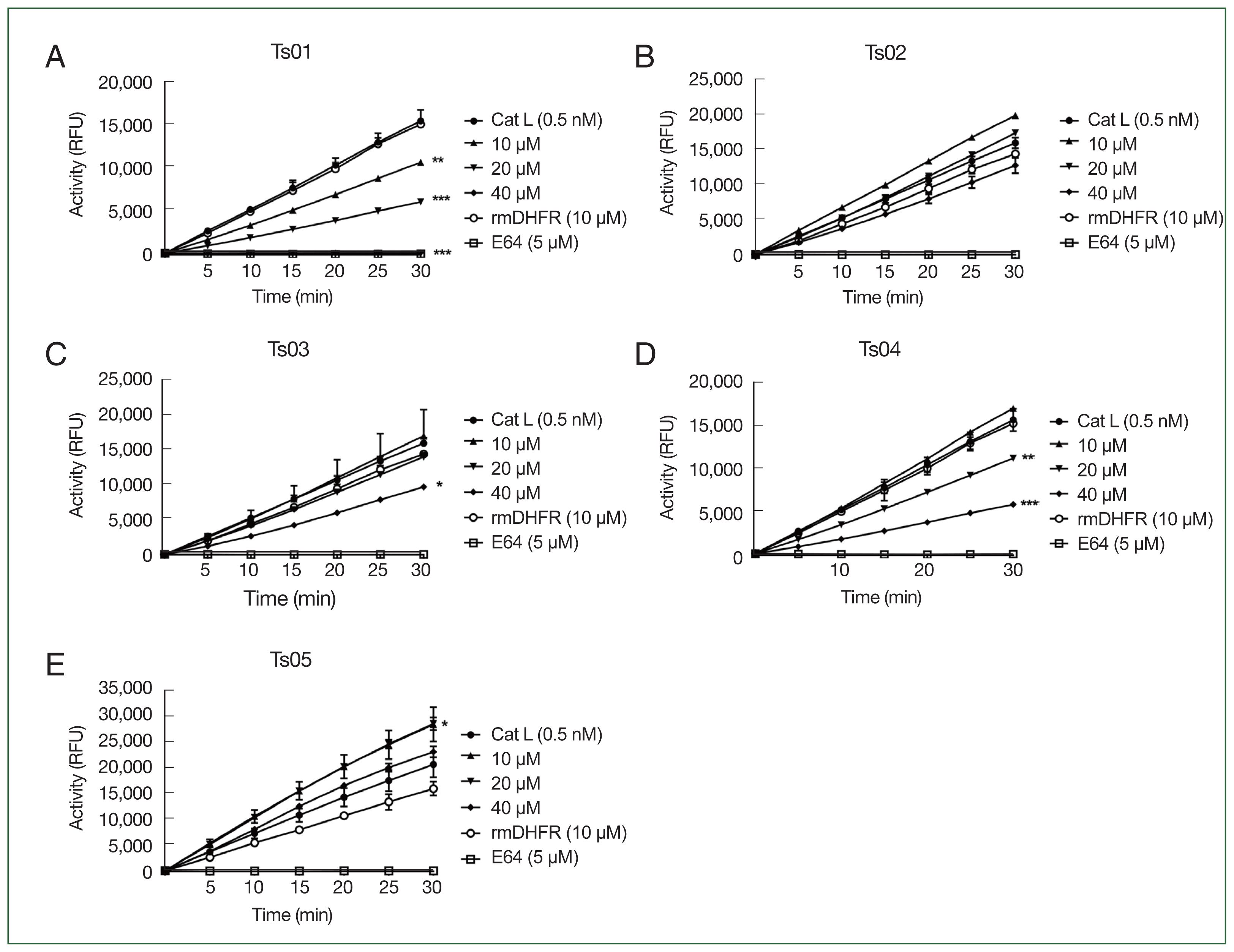
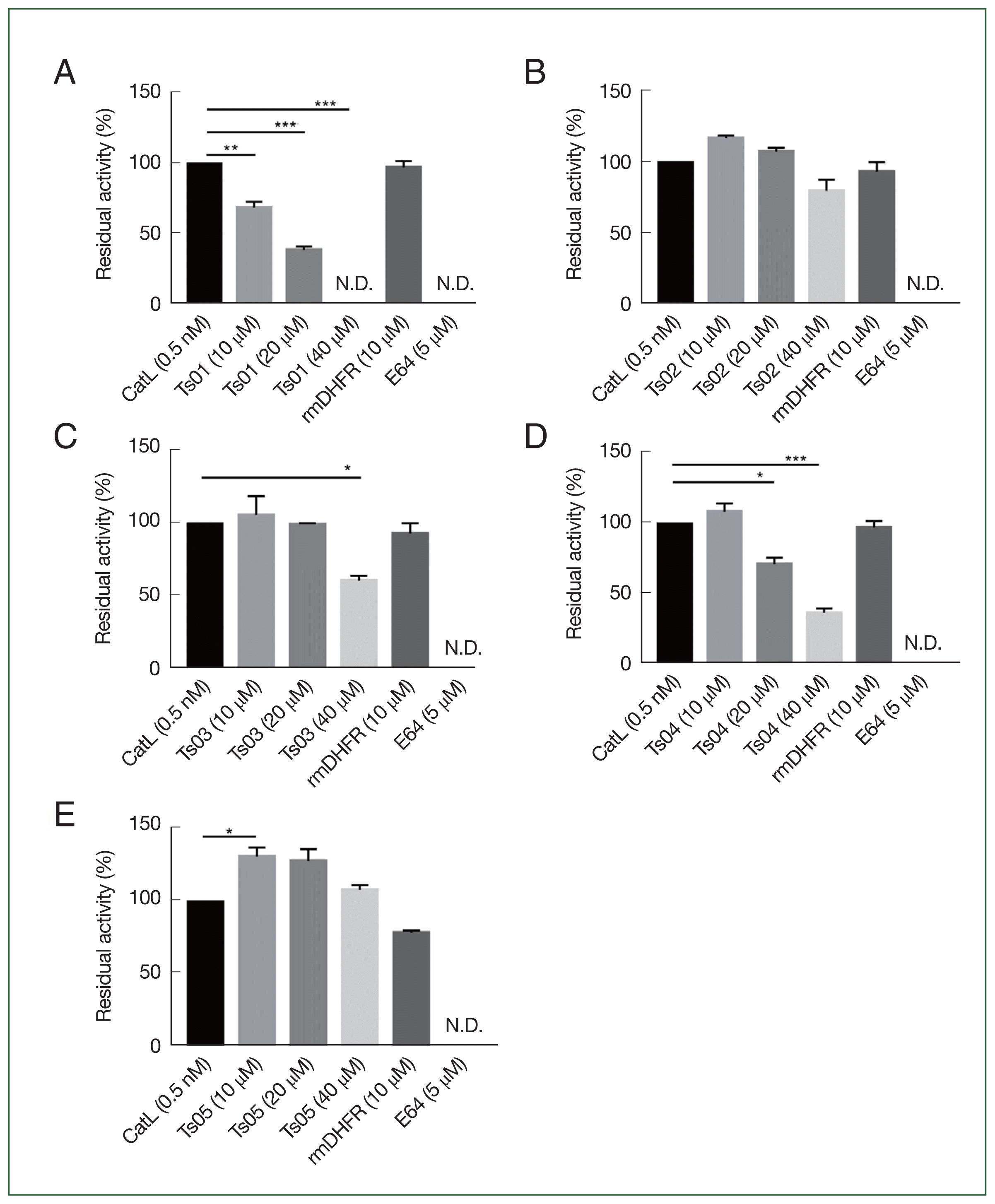
To evaluate the inhibitory properties of TsCstN truncates against CatL, different concentrations of rTs01 and truncates (10, 20, and 40 μM) were incubated with 0.5 nM CatL before reacting with a CatL-specific fluorogenic substrate. rTs01 significantly reduced CatL activity at concentrations of 10 μM (P<0.001), 20 μM (P<0.0001), and 40 μM (P<0.0001), with complete inhibition observed at 40 μM compared with CatL alone (Fig. 2A). Similarly, rTs04 significantly reduced CatL activity at 20 μM (P<0.001) and 40 μM (P<0.0001) compared with CatL alone (Fig. 2D), whereas rTs03 only inhibited CatL activity at 40 μM (P<0.05) compared with CatL alone (Fig. 2C). CatL activity was not inhibited by rTs02 (Fig. 2B). Surprisingly, rTs05 did not inhibit protease activity of CatL but did enhance protease activity (Fig. 2E). An irrelevant control protein (rmDHFR) did not suppress CatL activity while E64, a broad-spectrum cysteine protease inhibitor, completely inhibited CatL activity.
The percentage residual activity of CatL after treatment rTs01 and its truncates (at 10, 20, and 40 μM) at the endpoint is summarized in Fig. 3. These results align with the inhibitory effects observed in Fig. 2, where rTs01 and certain truncates demonstrated significant reductions in CatL activity.
Discussion
Cystatins derived from parasitic helminths have been characterized as immunomodulatory molecules that affect immune responses such as the suppression of T-cell proliferation, inhibition of antigen processing and presentation in antigen-presenting cells, and regulation of pattern recognition receptors, and particularly the TLR-4 receptor on macrophages and dendritic cells, which release suppressive cytokines enabling long-term survival in their host [21]. Our earlier research showed that TsCstN derived from T. spiralis reduced CatL-associated inflammation [6]. In this study, we focused on characterizing protease inhibitory regions of TsCstN to modify them into a small peptide inhibitor against excessive inflammation.
The sequence analysis of TsCstN identified a signal peptide (1–20 aa), an N-glycosylation region, and two potential disulfide bonds, which are indicated as type 2 cystatins. In our previous study, TsCstN was initially identified as a hypothetical protein, necessitating structural annotation to elucidate its potential function. The simulated 3D structure had the closest homology with human cystatin E/M (HsCstM). However, multiple alignment comparisons of the protein sequence of TsCstN with those of HsCstM and a few selected cystatin homologs through show that TsCstN lacks the conserved regions typical of type 2 cystatins. These include the absence of the glycine residue at the N-terminus, the QXVXG motif at L1, and the PW motif at L2 [6]. In the present study, additional helminth cystatin homologs were incorporated to ascertain the presence of cystatin-conserved regions. Despite these efforts, the results remained consistent with our earlier findings, and crystallization may be necessary to determine the accurate 3D structure of TsCstN. This would provide insights into the biological function of the protein and its significance in T. spiralis, including its role in host-parasite interactions. In previous studies, hypothetical proteins have been crystallized from parasites such as Giardia lamblia, Leishmania major, and Schistosoma mansoni [22–24]. This approach is crucial for gaining deeper insights into their biology and for the development of effective drugs and vaccines.
To clarify the inhibitory reactive region of TsCstN, the protein truncated sites were designed based on the 2D structure of TsCstN as predicted by five different programs, including Jpred4, PHD, Phyre2, PSIPRED, and PredictProtein. Impressively, all programs predicted a similar 2D structure of TsCstN, which comprised five reactive regions: α1, L1 (β1 and β2), irregular coil, L2 (β3 and β4), and α2. Four distinct truncates, Ts02–Ts05, were generated by deleting the protein regions in TsCstN. The 2D structures of all truncates, predicted by five different programs, showed consistent outcomes, indicating that the different deletions did not affect 2D structure conformation alterations.
I-TASSER and AlphaFold2 were used to simulate the 3D structures of Ts01 and truncates. AlphaFold2 could accurately predict the full-length and truncated TsCstN protein structures with a high degree of structural similarity to the hypothesized protein. Certain truncated structures, particularly Ts04, exhibited good quality with a high percentage of favored regions. I-TASSER is a widely used technique that creates precise 3D models of protein structures by combining several approaches, such as threading, ab initio modeling, and structural refining [17]. AlphaFold2 predicts the 3D structure of proteins by using a deep neural network architecture and advanced machine learning techniques that utilize amino acid sequences [18]. Consequently AlphaFold2 can predict 3D structures with high accuracy and prediction compared with I-TASSER. AlphaFold, an artificial intelligence-based technique for predicting protein structures, has demonstrated remarkable accuracy in model prediction and can consistently predict protein structures with atomic accuracy, even when no comparable structure is available [18]. Recently, AlphaFold has been extensively used in several structural biology fields, such as drug discovery, protein design, and protein function prediction [25–27].
The inhibitory properties of the recombinant TsCstN and truncates were confirmed in reactions against cathepsin L. Cysteine proteases, including CatB, CatL, and CatS, are involved in antigen presentation processing. CatB processes the antigen, and CatL and CatS degrade the major histocompatibility complex (MHC) invariant chain (li) in antigen processing cells, activating MHC-II formation [28,29]. Several studies have shown that parasite cystatins can inhibit cathepsin during antigen presentation [21]. In our previous study, rTsCstN specifically inhibited the activity of CatL but not that of CatB or CatS [6]. Therefore, this study focused on determining the inhibitory function of truncates against CatL. The 3D structure of TsCstN and other cystatins contains the NH2-terminal α1, L1, and L2, structures which are necessary to interact with the active site of cysteine proteases for inhibition [6,30]. rTs01 and rTs04 effectively inhibited CatL compared with other truncates. Thus, α1 and L1 are essential in inhibiting CatL activity, although rTs04 was less effective in reducing CatL activity than TsCstN-WT at the same dose. In human cystatin A, the NH2-terminal α1 region is involved in cysteine protease activity, and different NH2-terminal truncations cause the loss of inhibitory activity [31]. An important role of the α1 region of TsCstN in CatL inhibition was suggested by Ts02, which lacks α1 but contains L1 and L2. This truncate lost the ability to inhibit CatL. However, the α1 alone in Ts05 failed to inhibit CatL. Interestingly, rTs05 unexpectedly increased CatL activity instead of inhibiting it. However, the cause of this increase remains unknown and has not been addressed elsewhere and will therefore be explored in future study. We conclude that the α1 and L1 regions (Fig. 4), comprising 52 amino acid residues (DLSELDEAKNYIYQSDLQTGRGNFRKVLKVRNVDTSDGLSLTIDALPTTCPV) (Supplementary Table S3) are sufficient to inhibit CatL, whereas L2 helps increase CatL inhibitory activity.
Importantly, while TsCstN and its truncates exhibit CatL inhibitory activity, high concentrations of these recombinant proteins are required for effective inhibition. This could be due to several factors, including TsCstN not being a specific inhibitor for CatL or potential conformational inaccuracies when expressed in a prokaryotic system. Therefore, native TsCstN must be purified to test its protease inhibitory activity. Additionally, optimizing the expression system and conducting crystallization studies should be prioritized in future work.
In conclusion, CatL activity was efficiently inhibited by TsCstN truncates, particularly Ts04, which contains α1 and L1 regions. Further research should be conducted regarding the ex vivo reduction of macrophage inflammation and in vivo anti-inflammation in animal models. Furthermore, protein dynamics simulations, inhibitor types, enzyme kinetics, and molecular docking techniques can be used to deepen our understanding of the function of these proteins.