Morphological and molecular characteristics of Paralecithodendrium longiforme (Digenea: Lecithodendriidae) adults and cercariae from Chinese pipistrelle bats and viviparid snails in Thailand
Article information

Abstract
This study aimed to describe the morphological and molecular characteristics of Paralecithodendrium longiforme (Digenea: Lecithodendriidae) adults and cercariae isolated in Thailand. Adult flukes were isolated from the Chinese pipistrelle bat (Hypsugo sp.), and cercariae were detected in the viviparid snail (Filopaludina martensi martensi) from Chiang Mai province. The morphological characteristics were observed and described using conventional methods, and the molecular characteristics with internal transcribed spacer 2 (ITS2) and 28S rDNA gene sequences. The adult flukes were fusiform, 0.84–0.98 mm in length, and 0.37–0.49 mm in width, and were distinguishable from other species by the presence of longitudinal uterine coils. The cercariae were nonvirgulate xiphidiocercariae, with the oral sucker bigger than the acetabulum, the tail without fin fold, a body size of 117.5–138.3×48.3–52.2 μm, and a tail size of 100.7–103.7×15.0–18.9 μm. Molecular studies revealed that the adults and cercariae shared 99.3% (ITS2) and 99.6% (28S rDNA) homology with each other. They were phylogenetically close to P. longiforme with an identity of 94.5% for ITS2 and 98.7% for 28S rDNA. This study provides new information on the natural definitive host and first intermediate host of P. longiforme in Thailand. The discovery of its cercarial stage in Filopaludina snails highlights the importance of monitoring the associated second intermediate host and prevention and control of this potentially zoonotic trematode.
Introduction
Lecithodendriid flukes (Lecithodendriidae Lühe, 1901) are parasitic trematodes of fish, amphibians, mammals (mostly bats), and occasionally birds [1]. This trematode group comprises 12 genera namely, Vesperugidendrium, Castroia, Ophiosacculus, Gymnoacetabulum, Acanthatrium, Papillatrium, Glyptoporus, Caprimolgorchis, Ochoterenatrema, Lecithodendrium, Pycnoporus, and Paralecithodendrium [1]. Among these, two genera, Caprimolgorchis (C. molenkampi and Caprimolgorchis sp.) and Paralecithodendrium (P. glandulosum and P. obstusum), have been found in Thailand [2–4]. C. molenkampi (syn. Prosthodendrium molenkampi, Paralecithodendrium molenkampi) is a well-known species that infects humans in Thailand and Lao PDR [4].
Paralecithodendrium longiforme (Digenea: Lecithodendriidae) is a species of minute intestinal trematode that was first described as Lecithodendrium longiforme by Bhalerao [5] and isolated from the bat Nyctinomus plicatus in Myanmar. This trematode is distributed globally in various bat species from Africa, Europe, and Asia [6–9]. While P. longiforme is widely spread in many Asian countries, including Afghanistan, India, Japan, Malaysia, Vietnam, and the Philippines, it has not yet reported in Thailand. Despite numerous reports of this trematode in their definitive hosts, the life cycle of P. longiforme remained unexplained fully. According to previous studies, the life cycle of trematodes of the family Lecithodendriidae includes a prosobranch snail as the first intermediate host, aquatic insects as the second intermediate host, and vertebrates as the definitive host [10–13]. Notably, in prosobranch hosts, the cercarial stage of these trematodes has a stylet and is a virgulate or nonvirgulate xiphidiocercaria, which characterizes the larval stage of the family Lecithodendriidae [1]. The xiphidiocercariae have a high degree of morphological similarity with a variety of morphological features; however, identifying the larvae is difficult and requires a strong taxonomic background. Thus, a suitable molecular marker or gene region could be chosen to identify and deduce phylogenetic relationships [14,15].
The internal transcribed spacer 2 (ITS2) and 28S ribosomal DNA (28S rDNA) are considered highly conserved and have been used to study the developmental stages and to determine the phylogenetic relationships between trematodes in the family Lecithodendriidae. Kudlai et al. [16] described that the nucleotides of ITS2 and 28S rDNA of Paralecithodendium chilostomum (adults from the bat Nyctalus noctula in Ukraine) and Lecithodendrium linstowi (adults from the same bat species in Ukraine) matched well with high identity to Cercaria pugnax (an isolate from viviparid snails Viviparus viviparus in Ukraine) and Cercaria helvetica XII (an isolate from bithyniid snails Bithynia tentaculata in Lithuania), respectively. Enabulele et al. [17] reported that xiphidiocercariae found in the snail Radix balthica from the United Kingdom had 100% nucleotide similarity with L. linstowi adults from N. noctula in Ukraine and Pipistrellus pipistrellus bats in the UK based on ITS2 and large subunit (LSU) rDNA. In Thailand, Dunghungzin and Chontananarth [18] studied the phylogenetic tree of lecithodendriid xiphidiocercariae from two snail species (Bithynia siamensis and Filopaludina sumatrensis polygramma) based on the ITS2 region and revealed that the larvae were closely related to the family Lecithodendriidae (Lecithodendrium spp. and Mehraorchis spp.). No other molecular studies have been conducted on lecithodendriid flukes in Thailand. In this study, we aimed to describe the morphological and molecular characteristics of the adults and cercariae of P. longiforme recovered from the Chinese pipistrelle bat (Hypsugo sp.) and viviparid snails (Filopaludina martensi martensi) in Chiang Mai province, Thailand.
Materials and Methods
Ethics statement
All experimental hosts were managed according to the guidelines approved by the Animal Ethics Committee of the Faculty of Science, Chiang Mai University, Thailand, and the relevant document (no. RE004/19) was approved by the committee. This study complied with all the relevant national regulations and institutional policies for human care and use of animals.
Trematode detection in the host animals
The natural hosts (bats and snails) were investigated in a canal in Su Thep subdistrict, Mueang district (in Chiang Mai University), Chiang Mai province (18°48′15.6″N, 98°57′08.4″E) between February 2020 and January 2021 (Fig. 1). The sampling locations were surrounded by water resources and forests. Only one Chinese pipistrelle bat (Hypsugo sp.) was found dead near the canal at Chiang Mai University and was immediately transported to our laboratory. For the recovery of worms, the internal organs, including the intestines, were separated and thoroughly examined with the naked eye and under a stereomicroscope. A total of 35 viviparid snails (F. martensi martensi) were collected near the canal and transported to the laboratory. Each snail was crushed to detect larval trematodes, including xiphidiocercariae. The specimens were fixed in 4% formaldehyde solution on a glass slide for morphological studies and preserved in 95% ethanol in a sample tube for molecular studies. The prevalence of infection was assessed according to Bush et al. [19]. A total of 56 adult specimens of P. longiforme were collected from the intestine of the dead bat, and 3 out of 35 (8.6%) snails infected with the xiphidiocercariae of P. longiforme were examined.
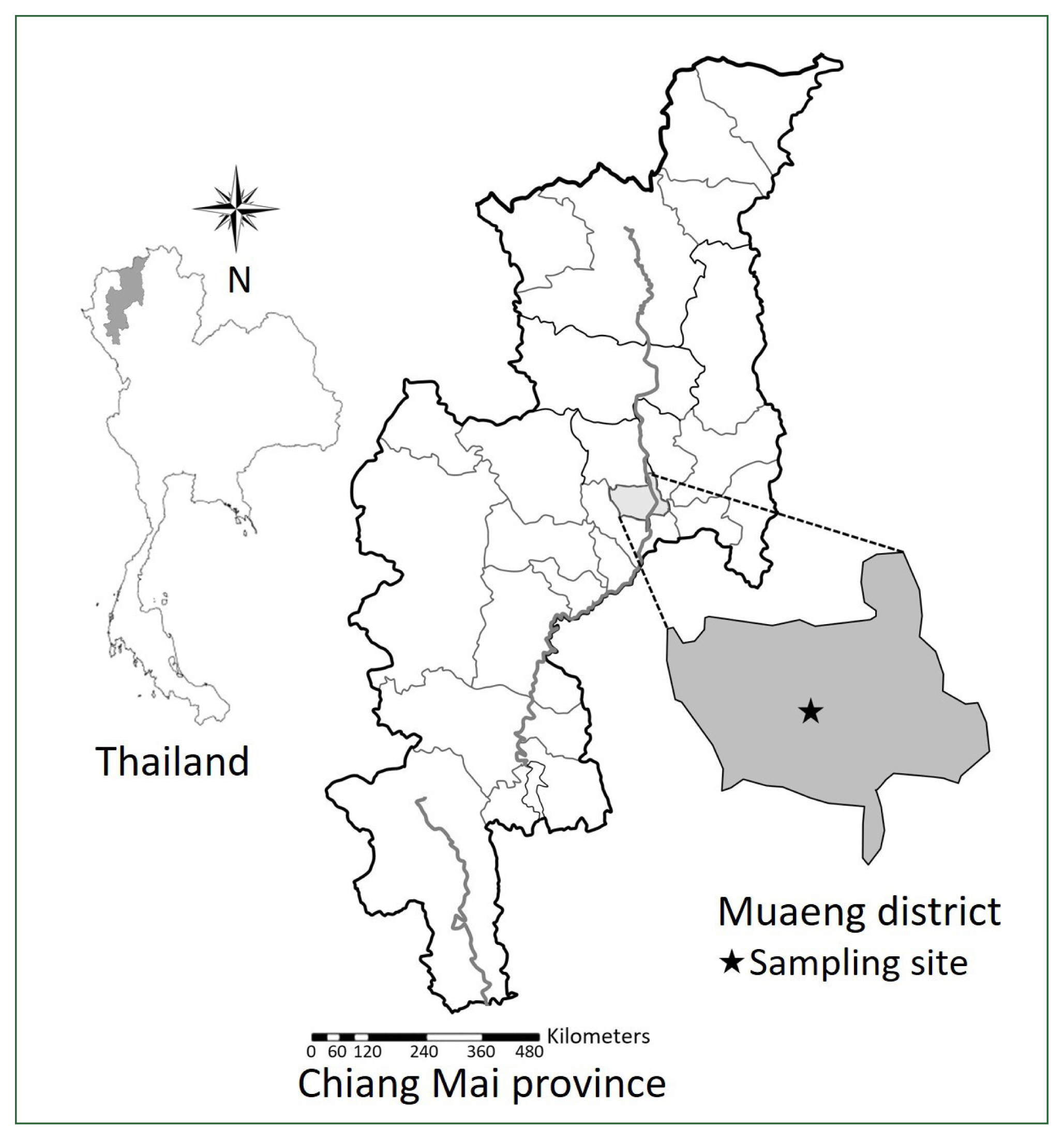
Sampling Iocation in Chiang Mai province, Thailand.
Morphological study
The specimens of adult and xiphidiocercariae were fixed with 4% formaldehyde between a glass slide and a cover slip overnight. The specimens were washed with distilled water and stained with Delafield’s hematoxylin (main stain) and eosin (counterstain). After staining with Delafield’s hematoxylin, the specimens were dehydrated with gradually increasing percentages of ethanol. If the stain was dark, it was removed from the specimen using a destaining solution (hydrochloric acid in 70% ethanol). The specimens were counterstained with a few drops of eosin in 85% ethanol. The specimens were cleared in xylene and transferred to a drop of Permount, which was placed on the glass slide and covered with a cover slip. The morphologies of the specimens were captured with a compound microscope using the software Optika Vision Pro (Optika, Bergamo, Italy) and drawn with the aid of a camera lucida attachment (Olympus, Tokyo, Japan). The muscular arrangement and nervous system were observed using a confocal laser scanning microscope equipped with Leica LAS X software (Leica, Wetzlar, Germany) and OlyVIA software (Olympus). The measurements of the specimens were designated by micrometers (μm) and millimeters (mm).
Molecular and phylogenetic analyses
The DNA of adult P. longiforme and xiphidiocercariae were individually extracted using Chelex solution [20] with 5% Chelex and 3 μl of proteinase K (Sigma-Aldrich, St. Louis, MO, USA). The specimens were heated at 55°C for 1 h, followed by vortexing and heating at 95°C for 30 min. The mixture was then centrifuged at 13,000 rpm for 1 min. The supernatant of each specimen was collected and preserved at −20°C until further use.
ITS2 region amplification was performed using the following primers: a forward 3S primer (5′-GGTACCGGTGGATCACTCGGCTCGTG-3′) and a reverse BD2 primer (5′-TATG CTTAAATTCAGCGGGT-3′) [21]. PCR was performed using the following cycling conditions: 94°C for 2 min, followed by 35 cycles of 94°C for 1 min and 60°C for 1 min, then 72°C for 1 min, and a final extension at 72°C for 7 min.
28S rDNA was amplified using 2 primers: a forward LSU-5 primer (5′-TAGGTCGACC CGCTGAAYTTAAGC A-3′) and a reverse 1500R primer (5′-GCTATCCTGAGGGAAACT TCG-3′), where Y represets C or T [22]. PCR was conducted using the following parameters: 94°C for 2 min, followed by 35 cycles of 94°C for 30 sec and 60°C for 30 sec, then 72°C for 75 sec, and a final extension at 72°C for 7 min.
The PCR products were separated by 1% agarose gel electrophoresis and stained with the nucleic acid stain, SafeViewTM FireRed (Applied Biological Materials, Richmond, British Columbia, Canada), and observed under ultraviolet light. The PCR products were cleaned and sequenced by Ward Medic Ltd., Part. (Bangkok, Thailand). The partial sequences were aligned and edited using MEGA version 10.1.8. The sequencing results were compared and analyzed using the BLAST program in NCBI (http://www.ncbi.nlm.nih.gov/blast/Blast.cgi). The phylogenetic trees were constructed using maximum likelihood (ML) and Bayesian inference (BI) methods from ITS2 and 28S rDNA datasets of relative species in the superfamily Microphalloidea. The outgroups included Centrocestus formosanus species (family, Heterophyidae). MEGA version 10.1.8 (Pennsylvania State University, USA) was used to run the ML trees, and node support was estimated using 1,000ML replicate bootstrapping of tree topologies [23]. Bootstrap values of 70 were treated as well supported [24]. MrBayes 3.1.2 was used to create the BI trees. Posterior probabilities <0.95 were deemed significant [25]. The ITS2 and 28S rDNA trees were analyzed using FigTree version 1.4.4. The ITS2 region and 28S rDNA gene sequences determined in this study are available in the NCBI database under the accession numbers OQ402582-83 (adults) and OQ402586-87 (cercariae).
Results
Morphological descriptions of adults
The shape of adult trematode (Figs. 2A, 3A; Table 1) was fusiform and moderate, measuring 0.84–0.98 mm×0.37–0.49 mm (n=10). The body tegument was smooth and without spines. The muscular oral sucker was well-developed and oval, located subterminally at the anteroventral side of the body, and measured 0.17–0.19 mm×0.14–0.15 mm. The prepharynx was short (0.01–0.02 mm in length) and was followed by the circular pharynx (Fig. 3A). The pharynx was 0.04–0.05 in diameter. The esophagus was longer than the prepharynx and was divided into two spacious saculiform intestinal ceca. The acetabulum was globular and well-developed, measured 0.08–0.09 mm in diameter and was situated at the middle line of the body. The acetabulum was smaller than the oral sucker. The pseudocirrus sac was circular, 0.04–0.05 mm in diameter, and was located between the acetabulum and ceca bifurcation area. The single ovary was slightly oval, measuring 0.05–0.06 mm×0.04–0.04 mm, and presented at the preacetabular area. Two circular testes were situated symmetrically at the level of the acetabulum and were 0.06–0.08 mm in diameter. The uterus occupied the entire posterior portion of the body. The vitelline follicle consisted of 20–24 follicles, which were anteriorly distributed to the split ceca on both sides of the body.
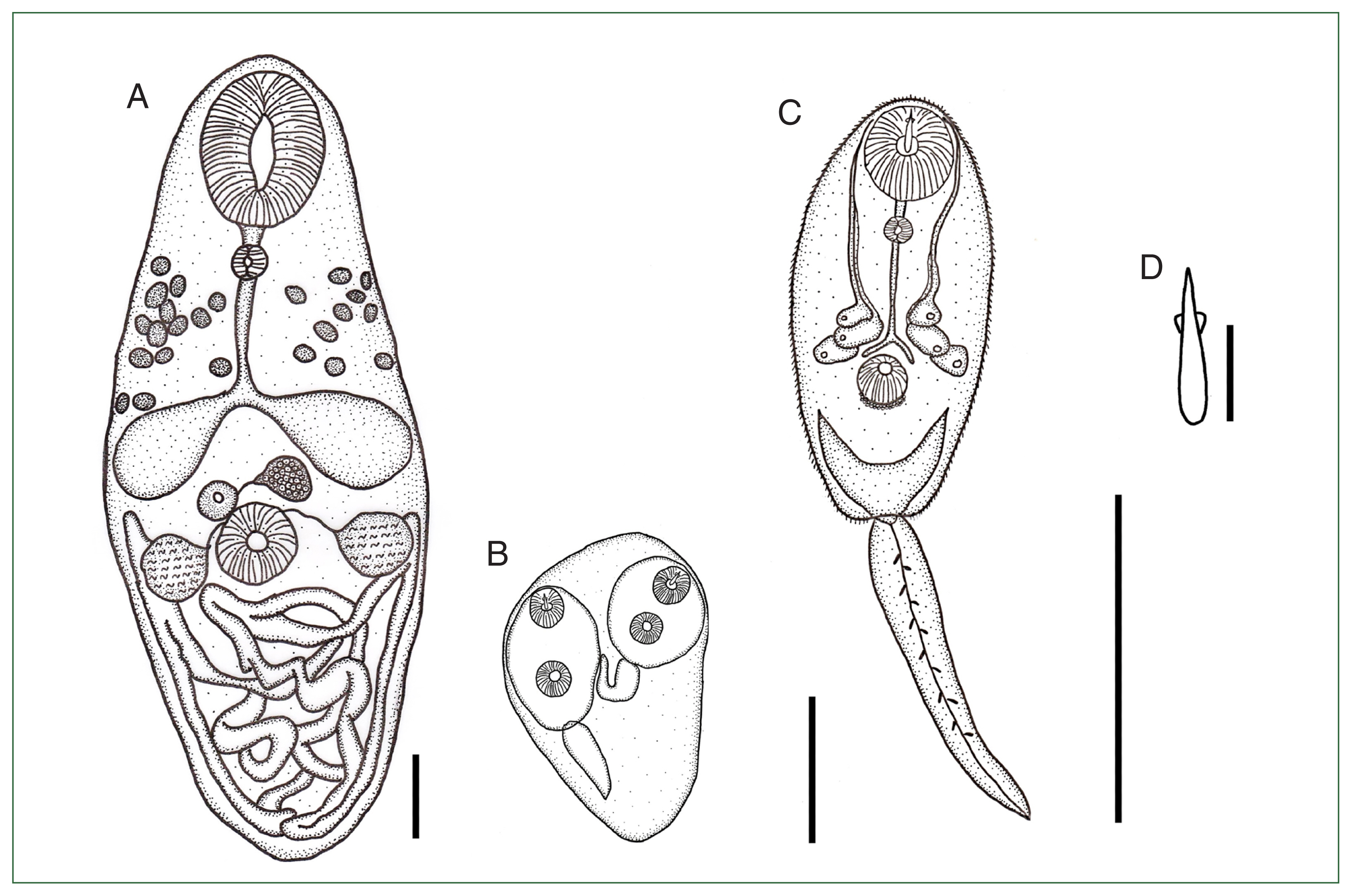
Illustration of Paralecithodendrium longiforme (adult) from the Chinese pipistrelle bat, Hypsugo sp. (A), sporocyst (B), and xiphidiocercaria (C) with the stylet (D) from the snail, Filopaludina martensi martensi from Chiang Mai province, Thailand. Scale bars: A=1 mm; B, C=0.1 mm; D=10 μm.
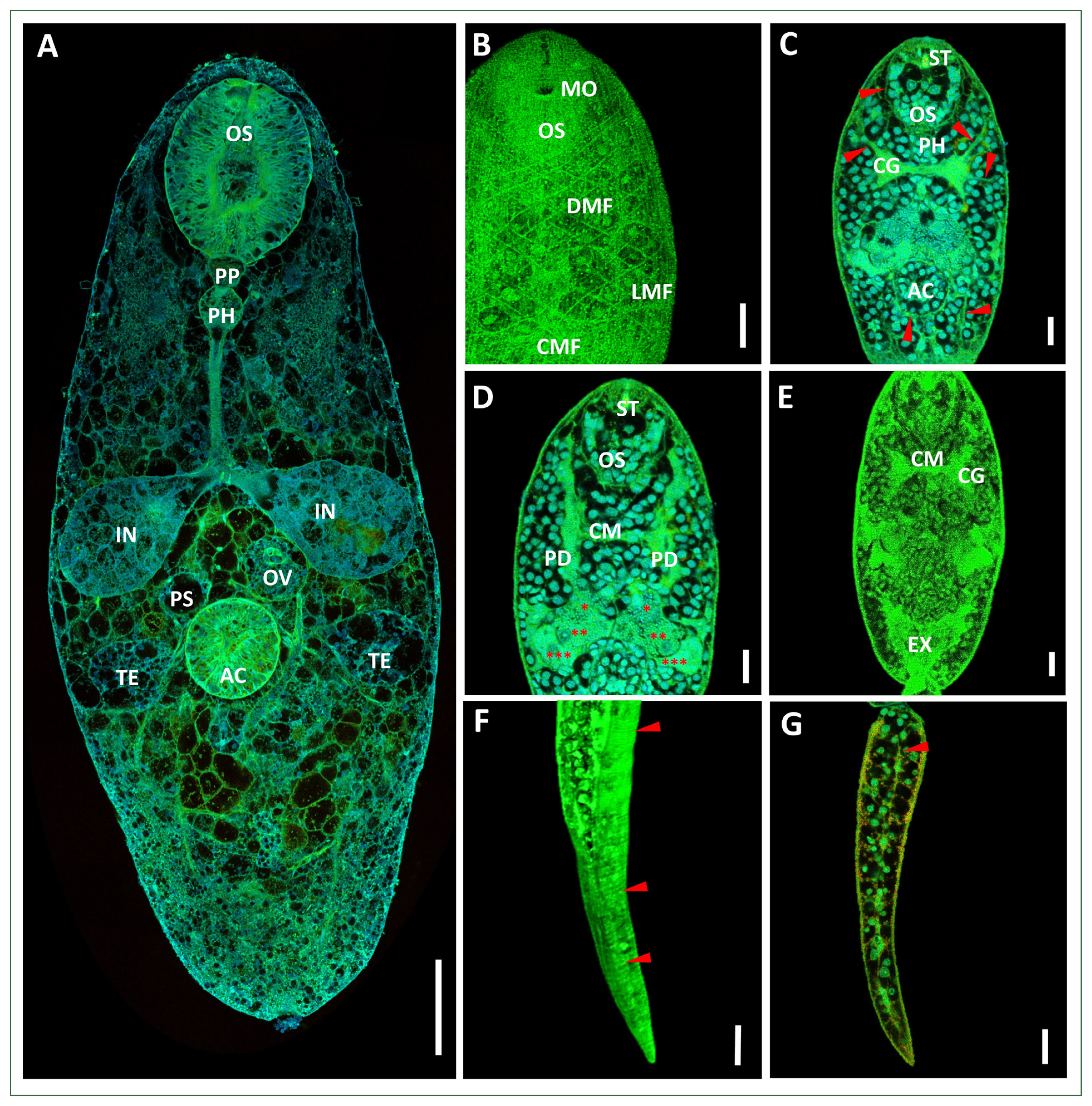
Confocal micrographs of the adult P. longiforme (A) and the xiphidiocercariae (B–G). (B) Musculature arrangement of the surface of the body. (C–E) Anterior body part with cephalic ganglions connected to the longitudinal nerve cord; the nerve cord extended toward the anterior and the posterior part of the body near the oral sucker and the acetabulum (red arrow). (D) Three pairs of penetration glands (red spots) connected with penetration ducts; each duct laterally extended upward and opened near the oral sucker. (E) V-shaped excretory bladder. (F) Annular transverse striation of the xiphidiocercaria tail (red arrow). (G) Excretory duct (red arrow). Scale bars: A=100 μm, B–G=10 μm. AC, acetabulum; CG, cephalic ganglion; CM, commissure; CMF, circular muscle fibers; DMF, diagonal muscle fibers; EX, excretory bladder; IN, intestinal caeca; LMF, longitudinal muscle fiber; MO, mouth opening; OV, ovary; OS, oral sucker; PD, penetration duct; PH, pharynx; PP, prepharynx; PS, pseudocirrus sac; ST, stylet; TE, testes.

Measurements (mm) of Paralecithodendrium longiforme adults (n=10) compared with previous studies
Morphological descriptions of sporocysts and xiphidiocercariae
The sporocyst (Fig. 2B) was an elongated oval. The tegument was thin and had no spines, measuring 124.6–297.0 μm in length and 113.0–157.9 μm in width (n=10). The matured sporocyst contained 1 to 6 xiphidiocercariae. A stylet was observed at the oral sucker of each xiphidiocercaria. The xiphidiocercaria (Figs. 2C, 3B–G) were developed in the sporocyst. The cercarial body was a small, elongate oval, measuring 117.5–138.3 μm×48.3–52.2 μm (n=10). The tegumental surface of the body was covered with tiny spines. The body wall consisted of an outer layer of circular, diagonal, and longitudinal muscle fibers. The layers of the circular muscle fibers were difficult to observe. The diagonal muscle fiber layers were conspicuously observed below the oral sucker and were widespread to the posterior of the body. The longitudinal muscle fibers were arranged at the extremities of the body and extended to the posterior end of the body. The diagonal muscle fibers were present with spaces wider than those of the longitudinal fibers (Fig. 3B). The well-developed oral sucker was situated at the subterminus, measuring 21.2–25.1 μm in diameter. The stylet was observed at the oral sucker, which was situated dorsally to the mouth opening. The stylet was lanceolate in shape, with a wide basal portion, and tapered upward, measuring 13.1–16.3 μm (Fig. 2D). The prepharynx was short, measuring 3.9–4.9 μm, and was followed by the pharynx. The pharynx was small and circular, measuring 4.7–6.0 μm in diameter. The esophagus was long, narrow, and split into two intestinal ceca at the anterior part of the acetabulum. Two cephalic ganglions occupied the anterior part of the body near the pharynx. The glands were connected by transverse commissures. Many nerve cords originated from the cephalic ganglions and extended toward the anterior and posterior parts of the body. Nerve cords were observed surrounding the oral sucker and the acetabulum (Fig. 3C). The well-developed acetabulum was 14.66–15.50 μm in diameter, which was smaller than the oral sucker. Three pairs of penetration glands were observed in the middle of the body, between the area of the pharynx and the split ceca. Each gland was connected to the penetration duct and laterally extended to the oral sucker area (Fig. 3D). The genital primordium was represented posterior to the acetabulum. The excretory bladder was V-shaped (Fig. 3E). The bladder was continued with the excretory canal of the tail and opened at the tail extremity. The tail was simple and flattened, measuring 100.7–103.7 μm×15.0–18.9 μm. The tail surface was covered with annular transverse striation (Fig. 3G). The excretory duct was also observed in the tail (Fig. 3G).
Molecular findings and phylogenetic tree
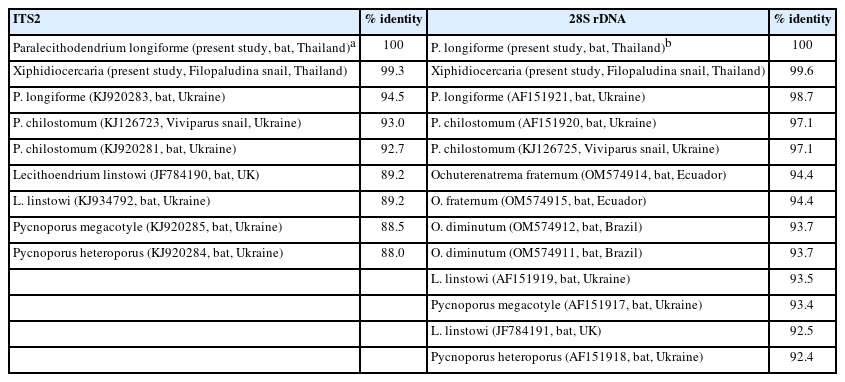
The ITS2 sequences of adults and xiphidiocercariae were successfully amplified for 399 bp, whereas the fragment of 28S rDNA obtained was 1,238 bp. The ITS2 tree (Fig. 4) revealed four families in the superfamily Microphalloidea: Lecithodendriidae, Collyriclidae, Prosthogonimidae, and Microphallidae. The lecithodendriid clade was separated into three genera: Paralecithodendrium, Lecithodendrium, and Pycnoporus. In this clade, P. longiforme adults isolated from the Chinese pipistrelle bat were clustered together with P. longiforme obtained from a bat (Myotis daubentoni) in Ukraine (identity: 94.5%) with a robustly supported clade (98ML/1BI). The xiphidiocercariae isolated from the snail F. martensi martensi were grouped with P. longiforme adults from this study (identity: 99.3%) with a high support clade (95ML/1BI) (Table 2).

Phylogenetic analysis of P. longiforme adults and xiphidiocercariae from this study and related species in the superfamily Microphalloidea focused on sequences of the ITS2 region. Centrocestus formosanus served as the outgroup. Numerical values at the branches represent nodal support by maximum likelihood (ML) bootstrap and Bayesian posterior probabilities (BI), respectively. ML values ≥70% and BI values ≥0.95 are shown. The new sequences are indicated in bold.
Sequence comparison of our specimens with members of the family Lecithodendriidae based on ITS2 and 28S rDNA
The 28S rDNA tree (Fig. 5) demonstrated five families of the superfamily Microphalloidea: Lecithodendriidae, Phaneropsolidae, Pleurogenidae, Prosthogonimidae, and Microphallidae. From the tree, the lecithodendriid clade was monophyletic. The family Lecithodendriidae represented four genera: Paralecithodendrium, Lecithodendrium, Pycnoporus, and Ochoterenatrema. P. longiforme from the present study was clustered together with P. longiforme obtained from the bat in Ukraine (identity: 98.7%) with a robust support clade (99ML/1BI). The xiphidiocercariae obtained in this study also appeared in the P. longiforme clade (99.6% identity with adults obtained in this study) with a high support clade (100ML/1BI) (Table 2).

Phylogenetic analysis of P. longiforme adults and xiphidiocercariae from this study and related species in the superfamily Microphalloidea focused on sequences of 28S rDNA. Centrocestus formosanus served as the outgroup. Numerical values at the branches present nodal support by maximum likelihood (ML) bootstrap and Bayesian posterior probabilities (BI), respectively. ML values ≥70% and BI values ≥0.95 are shown. The new sequences are indicated in bold.
Discussion
The morphological characteristics of P. longiforme observed in the present study fit the criteria of the genus Paralecithodendrium described by Lotz and Font [1]: the testes were located in the acetabulum level, which is close to the ceca ends, and the vitelline follicle was situated in the testicular or pretesticular level and lacks a pseudogonotyl. The morphologies and measurements of our specimens conformed to previous reports of P. longiforme from other countries (Table 1). Similarity in the longitudinal axis of the oral sucker, short saccular ceca, and the uterus entirely occupied the posterior half of the body was observed. The body length and width of our specimens overlapped with the specimens from Egypt [26] and Hungary [27]. The number of vitelline glands in this study was lower than reported by Bhalerao [5] and Pande [28], but were almost consistent with those reported by Saoud and Ramadan from Egypt [26]. Additionally, our specimens exhibited a short prepharynx similar to P. longiforme allahabadi (syn. Lecithodendrium longiforme allahabadi), which parasitizes the bat Nycticeius kuhli in India [28]. Dubois [29] considered P. longiforme allahabadi a synonym of P. longiforme. Recently, P. longiforme was also found in the mouse-eared bat Myotis sp. in the Philippines [9]. The presence of adult P. longiforme has been documented in various bats, including those from the families Rhinopomatidae, Emballonuridae, Megadermatidae, Rhinolophidae, Vespertilionidae, and Molossidae [6–9]. The present study identified the Chinese pipistrelle bat Hypsugo sp. (Vespertilionidae) as a new vertebrate host for P. longiforme. To the best of our knowledge, this study reports the presence of this trematode species for the first time in Thailand. Other larval stages of this trematode have not been recorded previously.
The nucleotide sequences of the ITS2 region and 28S rDNA were used to elucidate the other developmental stages of P. longiforme. Both the ITS2 and 28S rDNA sequences confirmed that P. longiforme were the xiphidiocercariae shed from the snail F. martensi martensi. The nucleotide sequences of xiphidiocercariae and adult P. longiforme showed similarity of 99.3% for ITS2 and 99.6% for 28S rDNA. The xiphidiocercariae in this study were assigned to the Microcotylae xiphidiocercaria group [30] because their body length was less than 200 μm, no virgula organ at their oral sucker, the acetabulum was located postequatorial to the body, the oral sucker was larger than the acetabulum, and the tail lacked a fin fold and was similar in length to the body. The virgula organ was an important feature that distinguished lecithodendriid-like species in the family Lecithodendriidae [1]. The virgula organ is a structure that contains mucoid secretion that penetrates its second intermediate host [31]. However, our larval specimens revealed the presence of nonvirgulate xiphidiocercariae. This result was similar to the xiphidiocercariae of P. chilostomum and Lecithodendrium sp. found in Europe [16], and a member of the Lecithodendriidae found in South America [32]. These findings established that the xiphidiocercariae from F. martensi martensi belonged to P. longiforme. To the best of our knowledge, this is the first documented report of the first intermediate host of P. longiforme.
In the phylogenetic tree, both the ITS2 and 28s rDNA sequences of the P. longiforme from this study were closely related to the P. longiforme reported in Ukraine and were separated from other species in the genus Paralecithodendrium (Figs. 4, 5; Table 2). This finding agreed with those of Tkach et al. [33] and Kudlai et al. [16], who found that the adults of P. longiforme and P. chilostomum were phylogenetically closer to each other than to the other genera of the family Lecithodendriidae. This close relationship between P. longiforme and P. chilostomum was supported by many morphological characteristics, including the pseudocirrus sac being located between the acetabulum, two split ceca and a vitelline follicle situated between the oral sucker and testes, and testes presented at the same height as the acetabulum [33]. However, P. longiforme is characterized by longitudinal uterine coils and an excretory bladder [5]. In addition, the xiphidiocercariae of P. longiforme and P. chilostomum appear to differ in their morphological characteristics; for example, the number of penetration glands (P. longiforme has three pairs whereas P. chilostomum has two pairs) and the stylet length (13.1–16.3 μm for P. longiforme vs. 28–30 μm for P. chilostomum) [16].
In Thailand, the metacercariae of lecithodendriid trematodes have been found in insects of the order Odonata, such as Brachythemis contaminata, Crocothemis servilia, Orthetrum sabina, and Trithemis pallidinervis, from northeast Thailand [2]. It was believed that the P. longiforme life cycle included insects as the second intermediate host because of the close systematic relationship at the family level. Hence, future surveys of metacercariae are warranted to determine the insect’s completion of the life cycle of P. longiforme.
In conclusion, we report for the first time P. longiforme adults and cercariae in Thailand and established their hosts: the Chinese pipistrelle bat Hypsugo sp. for adults and viviparid snails F. martensi martensi for the cercariae. This study elaborates on the geographic distribution and molecular data of a potentially zoonotic trematode parasite in the family Lecithodendriidae.
Acknowledgments
This research was supported by Chiang Mai University. The authors acknowledge the Applied Parasitology Research Laboratory, Department of Biology, Faculty of Science, Chiang Mai University for their facilities and laboratory processing.
Notes
Conceptualization: Arttra T, Butboonchoo P
Data curation: Arttra T, Wongsawad P, Wongsawad C, Nantarat N, Chai JY, Butboonchoo P
Formal analysis: Arttra T, Wongsawad P, Wongsawad C, Nantarat N, Chai JY, Butboonchoo P
Investigation: Arttra T, Wongsawad P, Wongsawad C, Nantarat N, Chai JY, Butboonchoo P
Methodology: Arttra T, Butboonchoo P
Project administration: Butboonchoo P
Validation: Arttra T, Wongsawad P, Wongsawad C, Nantarat N, Chai JY, Butboonchoo P
Writing – original draft: Arttra T, Butboonchoo P
Writing – review & editing: Arttra T, Chai JY, Butboonchoo P
We have no conflicts of interest related to this study.