Abstract
Human taeniases had been not uncommon in the Republic of Korea (=Korea) until the 1980s. The prevalence decreased and a national survey in 2004 revealed no Taenia egg positive cases. However, a subsequent national survey in 2012 showed 0.04% (10 cases) prevalence of Taenia spp. eggs suggesting its resurgence in Korea. We recently encountered 4 cases of Taenia saginata infection who had symptoms of taeniasis that included discharge of proglottids. We obtained several proglottids from each case. Because the morphological features of T. saginata are almost indistinguishable from those of Taenia asiatica, molecular analyses using the PCR-RFLP and DNA sequencing of the cytochrome c oxidase subunit 1 (cox1) were performed to identify the species. The PCR-RFLP patterns of all of the 4 specimens were consistent with T. saginata, and the cox1 gene sequence showed 99.8-100% identity with that of T. saginata reported previously from Korea, Japan, China, and Cambodia. All of the 4 patients had the history of travel abroad but its relation with contracting taeniasis was unclear. Our findings may suggest resurgence of T. saginata infection among people in Korea.
-
Key words: Taenia saginata, case report, molecular diagnosis, cox1, PCR-RFLP, sequence divergence
INTRODUCTION
Human taeniasis results from an intestinal infection with the parasitic tapeworm of the genus
Taenia. The disease is caused by 3 species including
Taenia asiatica, Taenia saginata, and
Taenia solium [
1]. Taeniasis is zoonotic because they involve pigs (
T. solium and
T. asiatica) or cattle (
T. saginata) as the intermediate host and humans as the definitive host [
2]. In case of
T. saginata, when cattle digest the eggs, oncospheres hatch and then invade the intestinal mucosa and migrate via circulation to muscles where they develop into metacestodes (=cysticerci). Humans can become infected by ingesting raw or undercooked infected beef [
2]. Usually in humans, a single
T. saginata worm is infected, grows to become an adult in about 3 months. The adult tapeworm produces gravid proglottids, which crawl out of the anus of the infected humans or are diffused in the feces [
2]. The main symptom is anal pruritis caused by outward migration of proglottids. Abdominal discomfort, mild diarrhea, and loss of body weight can also occur due to the presence of the tapeworm in the intestine [
2].
Before 1993, when
T. asiatica was first reported as a new species in the Republic of Korea (=Korea) [
3], all
T. asiatica had been considered as
T. saginata because of the almost indistinguishable morphology of the 2 species [
3]. Therefore, human
Taenia tapeworms in Korea were diagnosed as either
T. solium or
T. saginata, or undetermined until 1993 [
4]. In 1993, it was reported that
T. asiatica could be distinguished from
T. saginata by the existence of the rostellum on its scolex, presence of small posterior protuberances in gravid proglottids, higher numbers of uterine twigs, and wart-like formations on the larval bladder surface [
3]. However, these distinctions are difficult to observe in each individual strobila, and both morphological and genetic analyses were required to clearly distinguish the 2 species [
5].
Until the 1980s, in Korea, human taeniases had been one of the not uncommon parasitic infections [
6]. An old report in 1924 documented that the prevalence of
Taenia spp. eggs was 12.8%, which was the highest value of
Taenia prevalence ever recorded [
7]. After 1927, the prevalence declined gradually until it became 0% in a national survey of 20,541 randomly selected Korean people in 2004 [
1]. However, in a subsequent national survey performed in 2012 which targeted 23,956 people, 0.04% (10 egg-positive cases) prevalence of
Taenia spp.was detected [
8]. It was suggested that human taeniasis might be resurging recently.
In this article, we report 4 cases of T. saginata infection all of which were found during 6 months in 2013 (March to September), based on adult tapeworm recovery from the patients (neither T. asiatica nor T. solium cases during the same period). All the patients had the history of travel abroad to Asian countries, although no direct relationship was recognizable between the overseas travel and Taenia tapeworm infections. The specific identification of the worms was based on molecular analyses, including PCR-RFLP and partial sequencing of the cytochrome c oxidase subunit 1 (cox1) gene of the worms.
CASE RECORD
The 4 patients (cases A-D) found yellowish white tapeworm proglottids (4-5 in number in each patient) moving in their underwear or feces and consulted to the Department of Parasitology and Tropical Medicine, Seoul National University, Seoul, Korea. One of them complained of lower abdominal discomfort and anal itching; however, the other 3 had no special clinical complaints. All of them had experiences of visiting other countries including China, Cambodia, Japan, Dubai, or Turkey. Two of them tried flubendazole and albendazole based on wrong prescription, before prescribed with praziquantel in our department. All the 4 patients had the history of eating raw beef or rarely cooked beef steak recently (
Table 1).
In case B (48-year-old male), an almost complete strobila (about 1.1 m long) without scolex was obtained after treatment with praziquantel (15 mg/kg in a single dose) and purging with 40 g magnesium sulfate (MgSO
4). The gravid proglottids from the 4 patients revealed 16-20 lateral uterine branches and grossly looked like either
T. saginata or
T. asiatica. It was needed to do molecular analysis to obtain a specific diagnosis [
9-
11].
MATERIALS AND METHODS
We performed PCR-RFLP and gene sequencing of the mitochondrial
cox1 for specific identification of the 4
Taenia tapeworms. In order to use as positive controls, we obtained 3 known samples from the Parasite Resource Bank in Chungbuk National University (Cheongju, Chungbuk Province, Korea);
T. asiatica (cat. no. PRB081031800),
T. saginata (cat. no. PRB080110080), and
T. solium (cat. no. PRB071490001). The genomic DNA was extracted from a single segment by using the DNeasy® Blood & Tissue kit (Qiagen, Hilden, Germany) as recommended by the manufacturer. The mitochondrial
cox1 gene was targeted in PCR amplification. The PCR primers used were T1F (5'-ATA TTT ACT TTA GAT CAT AAG CGG-3') and T1R (5'-ACG AGA AAA TAT ATT AGT CAT AAA-3') [
9]. PCR was carried out in a 30 µl reaction mixture containing 15 µl of Smart 2x PCR Pre-Mix (SolGent Co., Ltd, Daejeon, Korea), 2 µl of template DNA, 10 µM of each primers, and 11 µl of distilled water. PCRs were progressed under 1 cycle of initial denaturation at 94℃ for 3 min followed by 40 cycles of denaturation (94℃ for 1 min), annealing (52℃ for 1 min), and extension (72℃ for 1 min), with a final extension at 72℃ for 10 min [
12]. For PCR-RFLP, the PCR products (10 µl) were digested with the restriction endonuclease (1 µl of
Nco1) in a final volume of 13 µl containing 2 µl of Ez-one 10x buffer for 1 hr at 37℃, and then analyzed by electrophoresis on 2% agarose gels.
The sequences of
cox1 from
Taenia tapeworm specimens were aligned using the Genieous program (version 6.1.4) (Biomatters Ltd., Auckland, New Zealand). The
Taenia tapeworms were identified based on the similarity of nucleotide sequences and phylogenetic relationships with those of
T. asiatica from Korea (GenBank no. AB465224), China (GenBank no. AB107235), and Japan (GenBank no. AB608742),
T. saginata from Korea (GenBank no. AB465246), China (GenBank no. AB107239), Cambodia (GenBank no. AB275143), and Japan (GenBank no. AB644391), and
T. solium from Korea (GenBank no. DQ089663). The neighbor-joining tree was constructed under the Kimura 2 parameter model by MEGA program version 5.2 to figure phylogenetic relationships [
13].
RESULTS
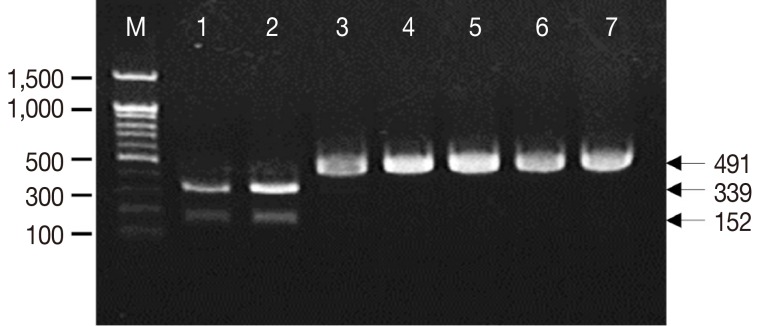
When the PCR products were digested with
Nco1 restriction enzyme, all the cases (A, B, C, and D) revealed identical PCR-RFLP patterns which were consistent with the known
T. saginata (
Fig. 1). Only 1 band (491 bp) was observed for
T. saginata, whereas 2 different bands (152 bp and 339 bp) were obtained for
T. solium and
T. asiatica.
Sequencing of the 491 bp
cox1 gene showed 99.8-100% identity with
T. saginata; however, only 94.7-94.9% and 87.8-88.0% identity was seen with
T. asiatica and
T. solium, respectively (
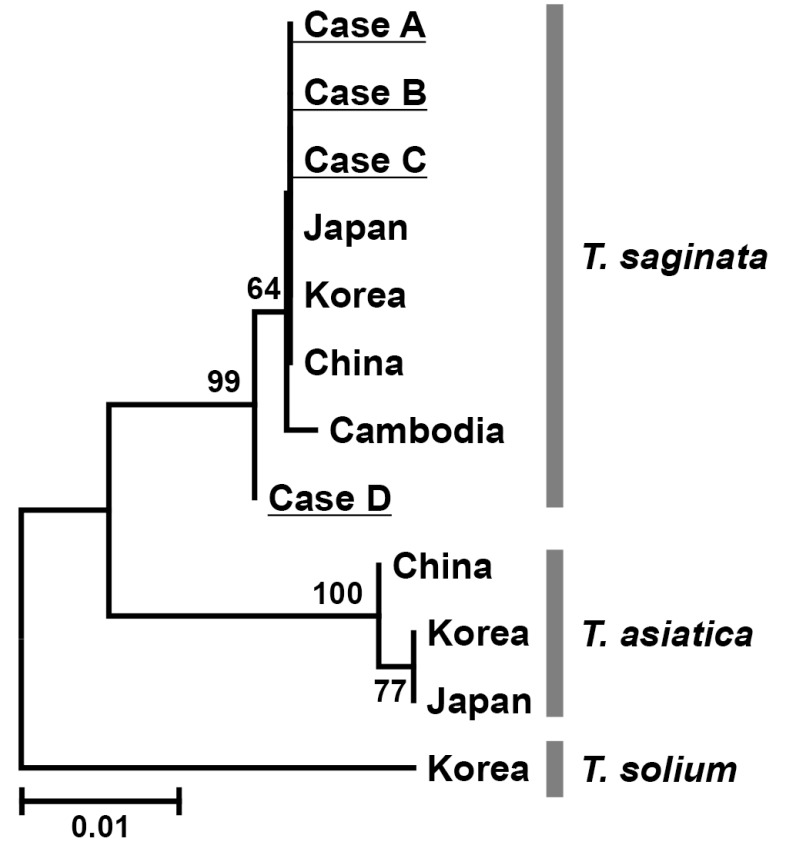
Table 2). Furthermore, the neighbor-joining tree revealed that our 4 specimens (cases A-D) were phylogenetically compatible to
T. saginata but far from
T. asiatica or
T. solium reported from various Asian countries and deposited in GenBank (
Fig. 2).
DISCUSSION
We used
Nco1 restriction enzyme, which recognizes and digests C^CATGG sites, to analyze the mitochondrial
cox1 gene of our
Taenia specimens. The
Nco1 was previously used for digestion of the
cox1 gene of
T. solium which was divided into 2 fragments [
14]. No other reports have been available which used this enzyme to digest
Taenia spp. genes. In the present study, we observed for the first time that
T. asiatica (from the Parasite Resource Bank, Chungbuk National University) was also divided into 2 fragments just like
T. solium (from the Parasite Resource Bank) both having the same recognition sequences by
Nco1. However, in case of
T. saginata (our specimens), there was no site for enzyme recognition by
Nco1, and thus the PCR products were preserved retaining only 1 band. Therefore, the
Nco1 enzyme appears to be useful to clarify the distinction between
T. saginata and
T. asiatica which are morphologically difficult to distinguish.
Korea had been one of the endemic areas for
Taenia spp. until the 1980s [
6]. However, several documents announced that the
Taenia egg positive rate became as low as 0.02% nation-widely in 1997 and finally dropped to 0% in 2004 [
15-
17]. Thereafter, only 5 sporadic cases were diagnosed during 2006-2011 [
1], and 10 additional cases were detected in 2012 in a national survey of 23,956 randomly selected people [
8]. With regard to the occurrence of
T. saginata, Jeon et al. [
6] reported 14 specimens of
T. saginata out of 68
Taenia spp. tapeworm specimens collected during 1935-2005 in Korea based on morphological and genetic analyses [
6]. After 2008, 2 cases of
T. saginata have been distinguished in Chungbuk National University [
1]. However, case reports with species identification of
T. saginata, including molecular analysis, have not been published since 2008. In this study, we reported 4 patients of
T. saginata infection who were referrred to our department during March-September 2013. All the 4 patients had the history of travel abroad but no evidence for contracting taeniasis abroad could be identified. No matter what the source of infection is, our findings may suggest resurgence of
T. saginata infection among the people in Korea.
Human
T. saginata infection is caused by ingesting raw or undercooked infected beef [
2]. However, in Korea, the infection status of cattle with
T. saginata metacestodes has seldom been documented except in a few old articles which described 17.8-21.3% prevalence in 1924 among cattle in Seoul and Gongju [
18], up to 37.6% in 1926 among cattle from different localities [
19], and 30.9% (1936) and 5.1% (1943) among cattle in Jeju-do [
20]. Particularly, after 1945, not a single official document has been published on the prevalence of
T. saginata metacestodes among the Korean cattle. Only one episode concerning a measled cattle (=infected with
T. saginata metacestodes) was available from a rural village of Gangwon-do in 1975 based on a personal communication [
4].
This study is significant to collect 4 cases of
T. saginata infection within a short time period of 6 months in a university parasitology department in 2013. Neither
T. asiatica nor
T. solium cases were detected during the same period in the department. Considering that only a limited number of
T. saginata cases occurred previously in Korea compared to
T. asiatica which occurred 4 times more frequently than
T. saginata [
6], there might be a resurgence of
T. saginata infection recently in Korea. Reactivation of the domestic life cycle of
T. saginata, if it was remained, and increased overseas travel of people, as well as import of beef from foreign countries, may be the possible responsible factors. Public attention should be paid to prevent human taeniasis avoiding intake of raw beef in Korea as well as in Asian countries endemic for taeniasis.
2012-0000037
Notes
-
We have no conflict of interest related with this study.
ACKNOWLEDGMENTS
Some parasite materials used in this study were provided by the Parasite Resource Bank of Korea, National Reserch Center (2012-0000037), Republic of Korea.
References
- 1. Chai JY. Human taeniasis in the Republic of Korea: hidden or gone? Korean J Parasitol 2013;51:9-17.
- 2. Dorny P, Praet N. Taenia saginata in Europe. Vet Parasitol 2007;149:22-24.
- 3. Eom KS, Rim HJ. Morphologic descriptions of Taenia asiatica sp. n. Korean J Parasitol 1993;31:1-6.
- 4. Eom KS, Rim HJ. Epidemiological understanding of Taenia tapeworm infections with special reference to Taenia asiatica in Korea. Korean J Parasitol 2001;39:267-283.
- 5. Eom KS. What is Asian Taenia? Parasitol Int 2006;55:S137-S141.
- 6. Jeon HK, Kim KH, Chai JY, Yang HJ, Rim HJ, Eom KS. Sympatric distribution of three human Taenia tapeworms collected between 1935 and 2005 in Korea. Korean J Parasitol 2008;46:235-241.
- 7. Uchida R. Results of fecal examination of intestinal helminthes among Korean prisoners in the Seodaemoon Jail. J Chosun Med Assoc 1924;46:74-83.
- 8. Korean Association of Health Promotion. Prevalence of intestinal parasitic infections in Korea. The 8th report. Seoul, Korea. 2013. p. 1-166 (monographic series in Korean).
- 9. Zarlenga DS, McManus DP, Fan PC, Cross JH. Characterization and detection of a newly described Asian taeniid using cloned ribosomal DNA fragments and sequence amplification by the polymerase chain reaction. Exp Parasitol 1991;72:174-183.
- 10. Bowles J, McManus DP. Genetic characterization of the Asian Taenia, a newly described taeniid cestode of humans. Am J Trop Med Hyg 1994;50:33-44.
- 11. Jeon HK, Eom KS. Taenia asiatica and Taenia saginata: genetic divergence estimated from their mitochondrial genomes. Exp parasitol 2006;113:58-61.
- 12. Jeon HK, Kim KH, Eom KS. Molecular identification of Taenia specimens after long-term preservation in formalin. Parasitol Int 2010;60:203-205.
- 13. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetic analysis using maximum likelihood, evolutionary distance, maximum parsimony methods. Mol Biol Evol 2011;28:2731-2739.
- 14. Margono SS, Ito A, Okamoto M, Subahar R, Yamasaki H, Hamid A, Wandra T, Purba WH, Nakaya K, Ito M, Craig PS, Suroso T. Taenia solium taeniasis/cysticercosis in Papua, Indonesia in 2001: detection of human worm carriers. J Helminthol 2003;77:39-42.
- 15. Korean Association of Health Promotion. Prevalence of intestinal parasitic infections in Korea. The 7th report. Seoul, Korea. 2004. p. 1-275 (monographic series in Korean).
- 16. Chai JY, Park JH, Guk SM, Kim HJ, Kim WH, Kim JL, Gu YS, Shin EH, Park HM, Hong KS, Kim SD, Lee SH. Status of intestinal parasite infections among 4,137 residents from provinces nationwide and metropolitan areas in the Republic of Korea (2004). Infect Chemother 2006;38:198-203.
- 17. Cho SH, Lee KY, Lee BC, Cho PY, Cheun HI, Hong ST, Sohn WM, Kim TS. Prevalence of clonorchiasis in southern endemic areas of Korea in 2006. Korean J Parasitol 2008;46:133-137.
- 18. Inoba J. Human taeniasis caused by eating meat in Chosen. Chou Jukai Zasshi 1924;37:229-239. (in Japanese).
- 19. Nakanish S. Statistical observation of Cysticercus inermis in native Korean calves. Nippon Jui Gakkai Zasshi 1926;5:147-150. (in Japanese).
- 20. Isshiki O. Intestinal helminth infections of domestic animals in Cheju. Chosen Gaku Ho 1960;16:1-13. (in Japanese).
- 21. Eom KS, Jeon HK, Rim HJ. Geographical distribution of Taenia asiatica and related species. Korean J Parasitol 2009;47:S115-S124.
Fig. 1Amplification of the cox1 mitochondrial gene of Taenia tapeworms by PCR-RFLP analysis. The amplified products of Taenia solium and Taenia asiatica were digested with Nco1 restriction endonuclease forming 2 different banding patterns (152 and 339 bp). Nonetheless, Taenia saginata and our 4 samples (491 bp) were not digested with the Nco1 enzyme. Lanes 1 to 7 are T. solium, T. asiatica, T. saginata (positive samples), and cases A, B, C, and D (test samples), respectively. M: 1 kb size maker (bp).
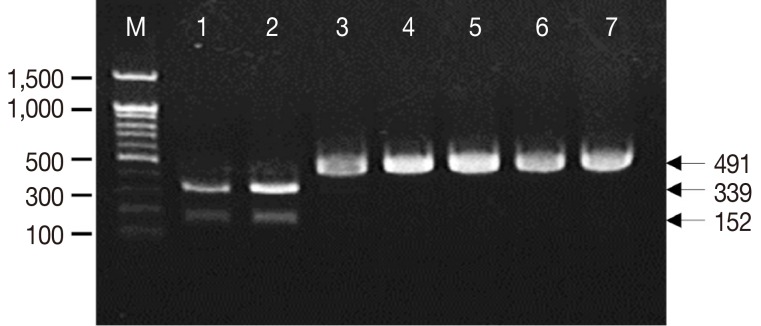
Fig. 2Neighbor-joining tree of human Taenia tapeworms based on nucleotide sequences of the cox1 gene. Numbers above the branches are bootstrap values. The scale bar represents the estimated number of nucleotide substitutions per nucleotide site. The phylogenetic tree reveals that the cox1 gene from our 4 patients (case A-D) was closer to T. saginata than T. asiatica or T. solium.
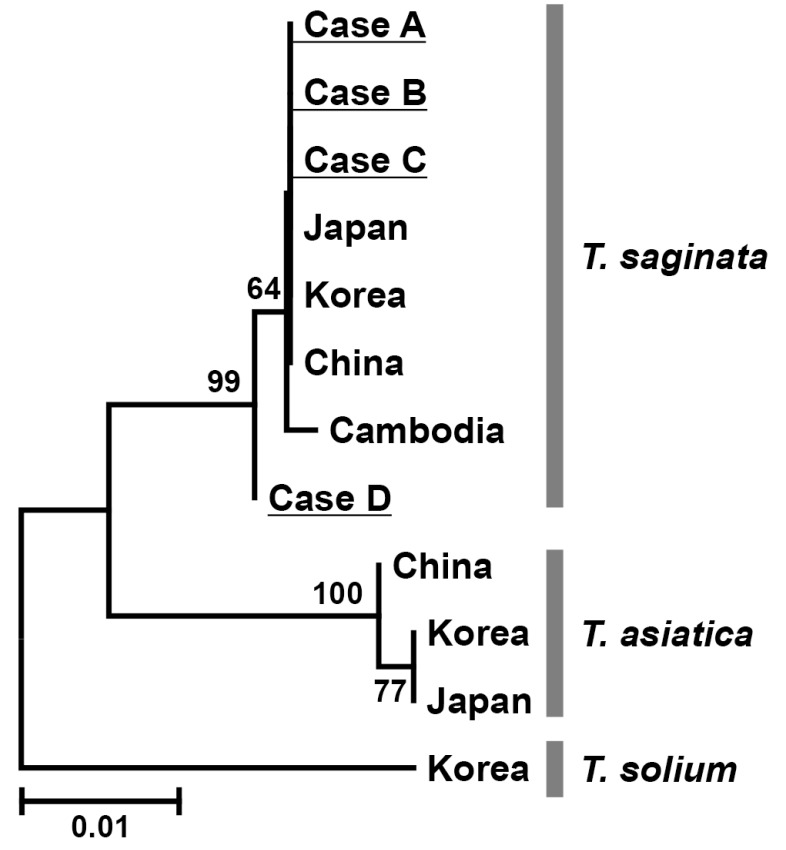
Table 1.Summary of 4 Taenia saginata cases
Table 1.
|
Case code |
Age & sex |
Clinical complaints |
Suspected source of infection |
Drug |
Visiting date to our department |
Countries traveled |
|
A |
27 M |
Passage of proglottids |
Raw beef |
Praziquantel |
Mar. 2013 |
China (2009) |
|
|
|
|
|
|
Cambodia (2010) |
|
B |
48 M |
Passage of proglottids, abdominal discomfort, anal itching |
Raw beef |
Praziquantel |
May-13 |
China (2004-2008) |
|
C |
30 F |
Passage of proglottids |
Rare beef steak |
Flubendazole |
Sep. 2013 |
Japan (2012) |
|
|
|
|
Praziquantel |
|
|
|
D |
36 M |
Passage of proglottids |
Raw beef |
Flubendazole |
Oct. 2013 |
Dubai (Sep. 2013) |
|
|
|
Rare beef |
Albendazole |
|
Turkey (Sep. 2013) |
|
|
|
sandwich |
Praziquantel |
|
|
Table 2.Nucleotide identity of our specimens with the known
Taenia species in GenBank
a for the mitochondrial cytochrome
c oxidase 1 (
cox1) gene
Table 2.
|
Our patients |
Taenia saginata
|
Taenia asiatica
|
Taenia solium
|
|
Case A |
491 (100.0) |
465 (94.7) |
431 (87.8) |
|
Case B |
491 (100.0) |
465 (94.7) |
431 (87.8) |
|
Case C |
491 (100.0) |
465 (94.7) |
431 (87.8) |
|
Case D |
490 (99.8) |
466 (94.9) |
432 (88.0) |