Abstract
Plasmodium vivax reemerged in 1993. It has been sustained for more than 25 years and become one of the important indigenous parasitic diseases in northern and western parts of the Republic of Korea near the demilitarized zone. In particular, relapse is a significant concern for the control of malaria, as short- and long-term incubation periods vary among those infected in Korea. In this study, the prevalence of asymptomatic carriers was examined among residents of high endemic areas of vivax malaria during nonseasonal transmission of mosquitoes. Blood samples from 3 endemic regions in northwestern Korea were evaluated by microscopic examination, rapid diagnostic testing, and nested PCR to identify asymptomatic patients carrying malaria parasites in the community. However, no positive malaria case among residents of endemic areas was detected. Additionally, serological analysis was carried out to measure antibodies against 3 antigenic recombinant proteins of P. vivax, merozoite surface protein 1-19, circumsporozoite surface protein-VK210, and liver-stage antigen (PvLSA-N), by the protein array method. Interestingly, seropositivity of sera between previous exposure and samples without exposure to malaria was significantly higher using the PvLSA-N antigen than the other antigens, suggesting that PvLSA-N can be used as a serological marker to analyze the degree of exposure for malaria transmission in endemic areas. This indicates a very low asymptomatic carrier prevalence during the nonmalaria season in the endemic areas of Korea.
-
Key words: Plasmodium vivax, diagnosis, resident, endemic area, non-malaria season, serological marker, PvLSA
INTRODUCTION
Malaria caused by
Plasmodium species of protozoan parasites is a major public health burden worldwide. In particular,
P. vivax is known to be less lethal than
P. falciparum, but it is more widely prevalent in tropical, subtropical, and temperate countries, including the Republic of Korea (Korea). Although eradication programs have been applied, vivax malaria has remained endemic to Korea since 1993, when the parasite was reemerged in a Korean Army soldier [
1,
2]. Unlike
P. falciparum, short- or long-term relapse of
P. vivax after sporozoite infection of hepatocytes can lead to subsequent infection without reinfection. Therefore, it is important to detect infection by the appropriate method and to treat patients with drugs targeting not only the blood stage but also the liver stage of
P. vivax [
3,
4]. In addition to relapse, asymptomatic patients infected with
P. vivax are occasionally the main factor in transmision of the parasite in the community; therefore, detection of asymptomatic patients is also essential for controlling and eliminating the parasite from humans.
Several advanced diagnosis techniques have been developed and applied. For example, microscopic examination with Giemsa-stained thick and thin blood films is still widely used as a gold standard method; it is rapid and inexpensive for detecting the parasite in blood samples. However, this method has the disadvantage of requiring expert specialists in field clinics and/or laboratories, a lack of which can lead to misdiagnosis [
5,
6]. For example,
P. knowlesi is morphologically undistinguishable from
P. malariae by microscopy, suggesting that multiple detection methods must be applied for detection of the malaria parasite.
PCR-based molecular diagnosis methods are relatively sensitive and accurate compared to microscopy and rapid diagnostic tests (RDTs) for malaria detection, and PCR can be applied for genus- or species-specific diagnosis, even in cases of low-level parasitemia not detected by RDTs or microscopic examination [
7]. In addition to molecular-based methods, serological diagnosis methods have been developed and applied for not only current infection but also previous exposure to the malaria parasite. Infected patients have specific antibodies against antigens expressed by the parasite, and these antibodies can be used as a serological marker. Accordingly, this method is useful for tracking the trend of transmission in a village and/or community of endemic areas for the purposes of surveillance [
8].
In the present study, residents living in endemic areas in Korea were recruited and analyzed for malaria infection by microscopy, PCR, and RDTs. In addition, we compared seropositivity to specific Plasmodium vivax antigens, namely, merozoite surface protein (PvMSP1), circumsporozoite protein (PVCSP) and liver-stage antigen (PvLSA), as recombinant proteins between healthy individuals and a pre-exposure group to identify the best serological marker available for assessing transmission status in high endemic areas of Korea.
MATERIAL AND METHODS
Sample collection and preparation
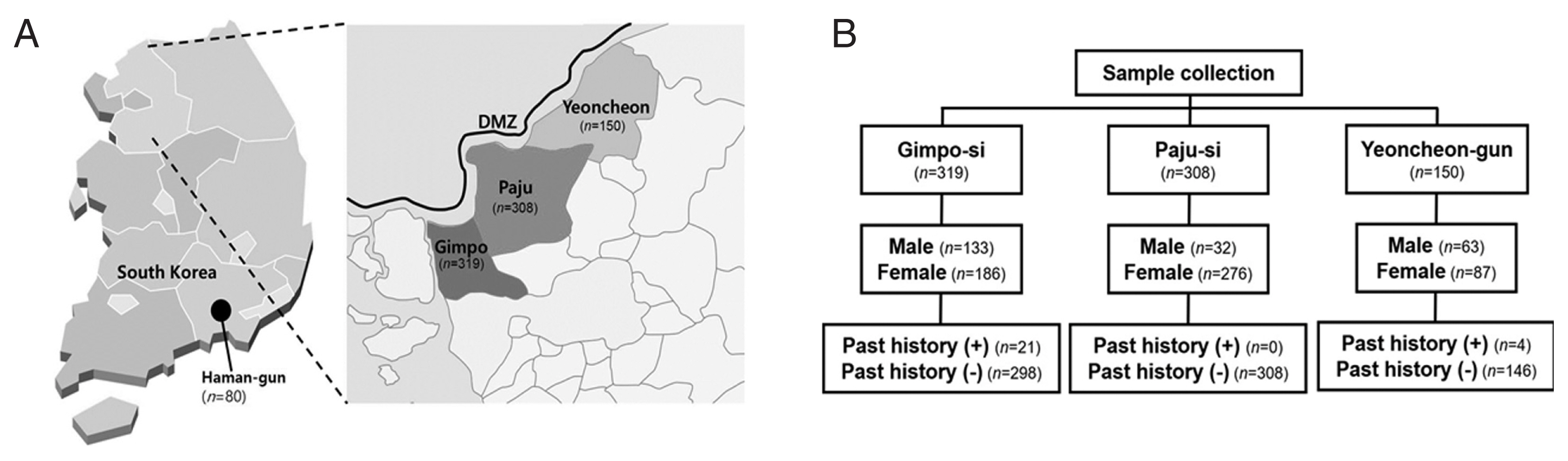
The blood samples used in this study were collected from residents in 3 regions on April 2019. A total of 777 blood samples from Gimpo-si (n=319), Paju-si (n=308), and Yeoncheon-gun (n=150) from local health centers, where malaria cases are diagnosed in a relatively higher number compared to other endemic areas from nation-wide surveillance of 2005 through 2018, were obtained (
Fig. 1A). In addition, we used serum samples from Haman-gun (n=80), Gyeongsangnam-do as vivax nonendemic areas for comparison of antibody response with those of endemic areas. Blood samples confirmed to be either vivax positive or negative by testing with a rapid diagnosis test kit and microscopy were collected in endemic areas of Korea. The blood samples collected in BD Vacutainer EDTA tubes (Becton Dickinson, Franklin Lakes, NewJersey, USA) were delivered to Kangwon National University; confirmation by blood films for microscopic examination and RDT was performed. The plasma and packed red blood cells were separated and stored at −80°C for protein array and genomic DNA isolation, respectively. The Institutional Review Board approved this study at Kangwon National University (no. 2019-03-002-001).
The pooling method for genomic DNA extraction was used, as described in our previous study [
9]. Briefly, 20 μl each of 10 different samples was pooled for extraction of genomic DNA using an AccuPrep Genomic DNA Extraction Kit (Bioneer Corp., Daejeon, Korea) in accordance with the manufacturer’s instructions. Nested PCR was performed as described in a previous report [
10]. Briefly, for the primary reaction, 4 μl of the eluted gDNA was mixed in a 20 μl reaction mixture using Accupower premix (Bioneer) with rPLU1_F (TCAAAGATTAAGCCATGCAAGTGA) and rPLU5_R (CCTGTTGTTGCCTTAAACTTC) primers. In the second nested PCR, 2 μl of the first amplicon was used as a template under the same 20 μl reaction mixture conditions as described above. rPLU3_F (TTTTTATAAGGATAACTACGGAAAAGCTGT) and rPLU4_R (TACCCGTCATAGCCATGTTAGGCCAATACC) were used for the nested PCR. The PCR conditions were as follows: step 1, 95°C for 5 min; step 2, denaturation at 95°C for 30 sec; step 3, annealing at 55°C for 1 min; step 4, extension at 72°C for 2 min in the first PCR and 30 sec in the second PCR. Steps 2 to 4 were repeated 30 times with the annealing temperature changed to 60°C, and step 4 was performed for 10 min. The amplified products were visualized on 1.5% agarose gels.
CareStartTM Malaria Pf/PAN (HRP2/pLDH) Ag Combo RDT (AccessBio Inc., Somerset, NewJersey, USA) was used with whole-blood samples from endemic areas according to the manufacturer’s manual. Blood samples from patients with vivax malaria were used as a positive control.
Cloning and recombinant protein expression
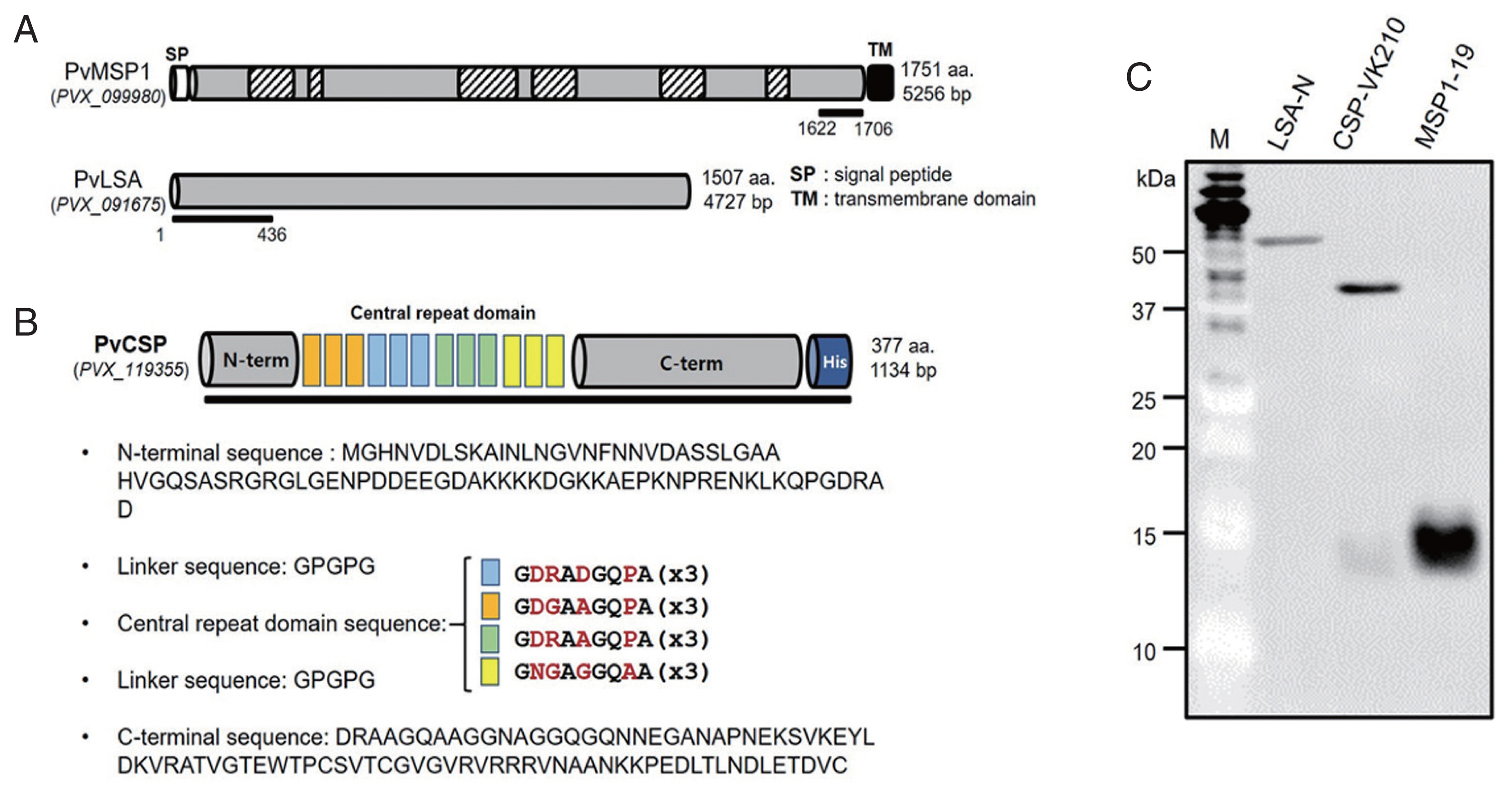
Genomic DNA extracted from the whole-blood sample of vivax malaria patients was used as a template for gene amplification of
pvmsp1-19 (GenBank accession no. XM_001614792.1;
Fig. 2A). The N-terminal region of
pvlsa (XM_001615278.1;
Fig. 3A) and full-length
pvcsp-
vk210 (XM_001613018.1;
Fig. 2B) were synthesized by Macrogen (Seoul, Korea). The central repeat domain of
pvcsp was specially designed from previous sequence analysis reports (
vk210 genotype mostly) of Korean vivax isolates. Segments of the genes were amplified using a high-fidelity KOD-plus Kit (TOYOBO Co., Osaka, Japan) with the following pairs of gene-specific primers:
pvmsp1-19 forward (gggcggatat
ctcgagAACGAGTCCAAGGAAATATTATCCCAG) and reverse (gcggtacccg
ggatccGTCCGTCATTTTACATTCAGCTTCAGG);
pvcsp-vk21 forward (gggcggatat
ctcgagATGAAGAACTTCATTCTCTTGGCTGTTTCTT) and reverse (gcggtacccg
ggatccCTAATTGAATAATGCTAGGACTAACAATATGACTAGC); and pvlsa-n forward (gggcggatat
ctcgagATGAGTTCCTTCTGGAGCTTCATCAAAT) and reverse (
gcggtacccgggatccCTAACTCCTCGAGATGTCATCGTTGCT). Small letters indicate the plasmid-derived sequences for in-fusion cloning, and
BamHI and
XhoI restriction enzyme sites are indicated as italicized and underlined letters. The amplicons were purified and cloned into the pEU-E01-His-Tev-N2 plasmid vector (CellFree Sciences, Matsuyama, Japan) designed for the wheat germ cell-free protein expression system by the In-fusion cloning method (Takara Bio USA Inc., Mountain View, California, USA). The sequences of cloned plasmids were confirmed. The recombinant proteins were synthesized by the wheat germ cell-free protein expression system using the bilayer translation reaction method, as described previously [
11]. The expressed recombinant proteins were confirmed by western blot analysis under reducing conditions using an anti-His-tagged antibody (Qaiagen, Hilden, Germany).
Amine-coated glass slides were prepared as described in our previous study [
12]. The crude recombinant protein antigens (1 μl) were spotted onto amine-coated glass slides in duplicate and incubated at 37°C for 2 hr. After blocking with 5% BSA in PBS containing 0.1% Tween-20, the recombinant protein-coated slides were reacted with each of the individual serum samples in duplicate. Fluorescence was detected with an Alexa Fluor 546-conjugated goat anti-human IgG secondary antibody (Invitrogen, Carlsbad, California, USA) and an Innopsys IS300 scanner (InnoScan, Carbonne, France). Mean fluorescence intensity (MFI) plus 2 standard deviations (2SD) was used as the cut-off value. The MFI was normalized by dividing the MFI by the cut-off value, and the normalized MFI from patients was subtracted from the normalized MFI from healthy serum.
Statistical analysis was performed using the Statistical Package for the Social Sciences (SPSS) version 16.0 (SPSS Inc., Chicago, Illinois, USA). The unpaired t-test was used for nonparametric independent 2-group comparisons between the healthy individual group and the vivax malaria patient group. Differences with P<0.05 were regarded as statistically significant. MFI values were plotted using GraphPad Software version 5.0 (GraphPad Software, La Jolla, California, USA).
RESULTS
Demographic characteristics of the study population and malaria diagnosis with microscopic examination, RDT, and PCR
In this study, a passive case detection method was carried out by local health authorities, and a total of 777 residents from 3 areas, Gimpo-si (n=319; age 70.8 on average and 93-31 on range), Paju-si (n=308; 70.5 and 95-23), and Yeoncheon-gun (n=153; 59.2 and 90-20), were recruited for the detection of clinical malaria (
Fig. 1). All collected blood samples were confirmed by blood smear and RDT (data not shown) as well as the nested PCR method (data not shown) for malaria infection. Consequently, there was no positive case based on the 3 methods; however, the amplified gene was observed in the positive control sample.
To analyze serological positivity for
P. vivax proteins, 3 antigens,
P. vivax merozoite surface protein 1 (PvMSP1),
P. vivax circumsporozoite protein (PvCSP), and P. vivax liver-stage antigen (PvLSA), were investigated (
Fig. 2A, B). The full-length C-terminal region of PvMSP1 (aa 1622-1706), including a combination of the central repeat region sequence of PvCSP (aa 1-377) and the N-terminal region of PvLSA (aa 1-436), was used for recombinant protein expression. The recombinant proteins were successfully expressed by the wheat germ cell-free expression system, and the proteins were detected by western blot analysis (
Fig. 2C).
The protein array method was carried out to analyze the seropositivity of the plasma samples collected from 777 participants as well as 16 vivax patients and healthy individuals as controls. The serum samples were diluted (1:25) and reacted with 3
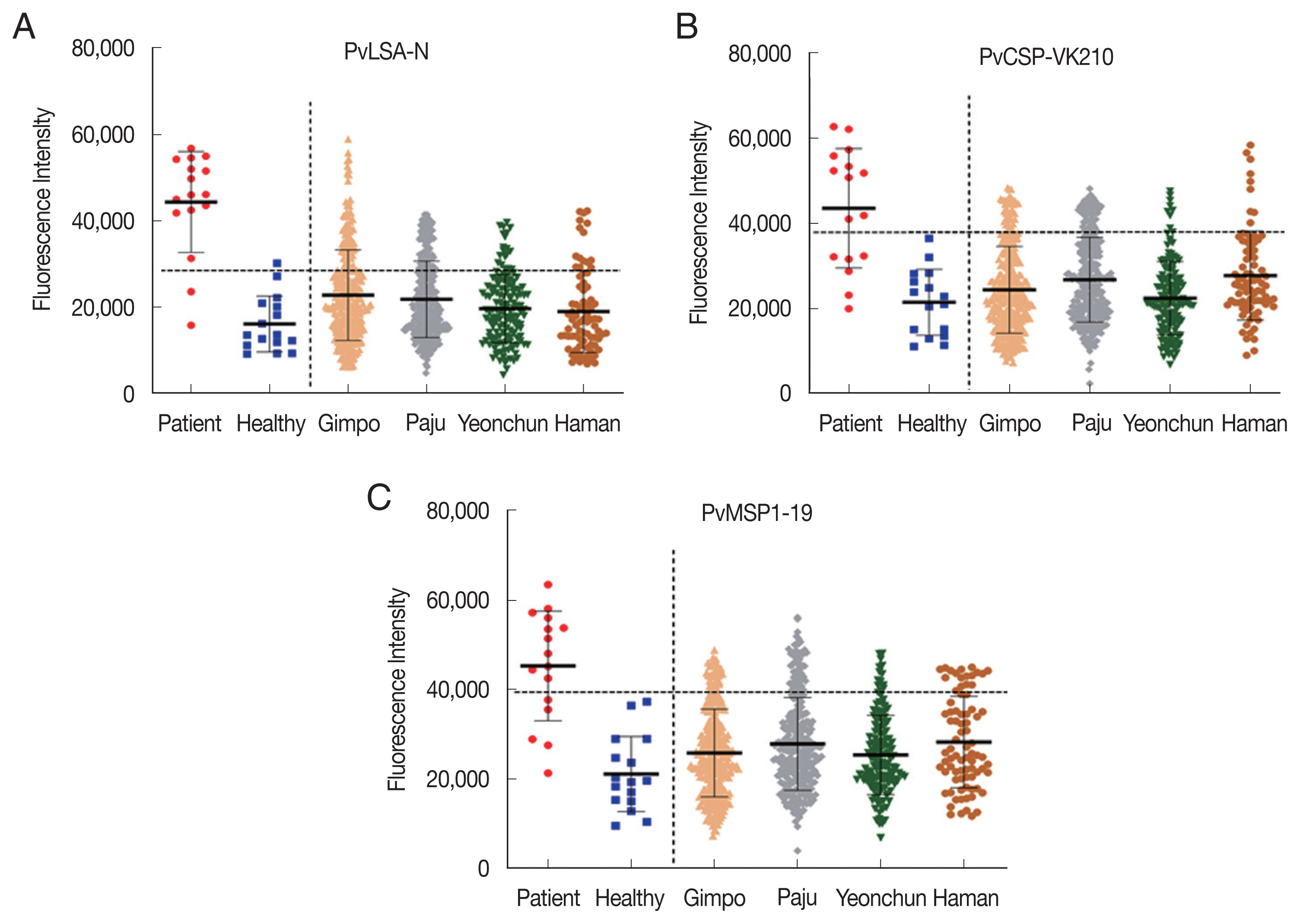
P. vivax antigens bound on array chips. For the control, PvLSA-N showed higher positivity to patient sera (87.5%) than PvCSP-VK210 (62.5%) and PvMSP1-19 (68.7%). However, one healthy serum sample was positive for PvLSA-N but not for the others (
Fig. 3;
Table 1). Among plasma collected from residents of 3 local surveyed areas, the seropositivities of PvLSA-N (20.2±6.8%) and PvCSP-VK210 (15.1±6.3%) were higher than 18.8% and, 16.3%, respectively, in Haman residents but not in cases of antibody titers against PvMSP1-19 (
Fig. 3;
Table 2).
We also issued questionnaires about previous exposure to malaria infection to the residents of the surveyed areas. Gimpo-si and Yeoncheon-gun had 21 and 4 cases of malaria history, respectively, with exposure to vivax infection from a few years to 50–60 years previously (
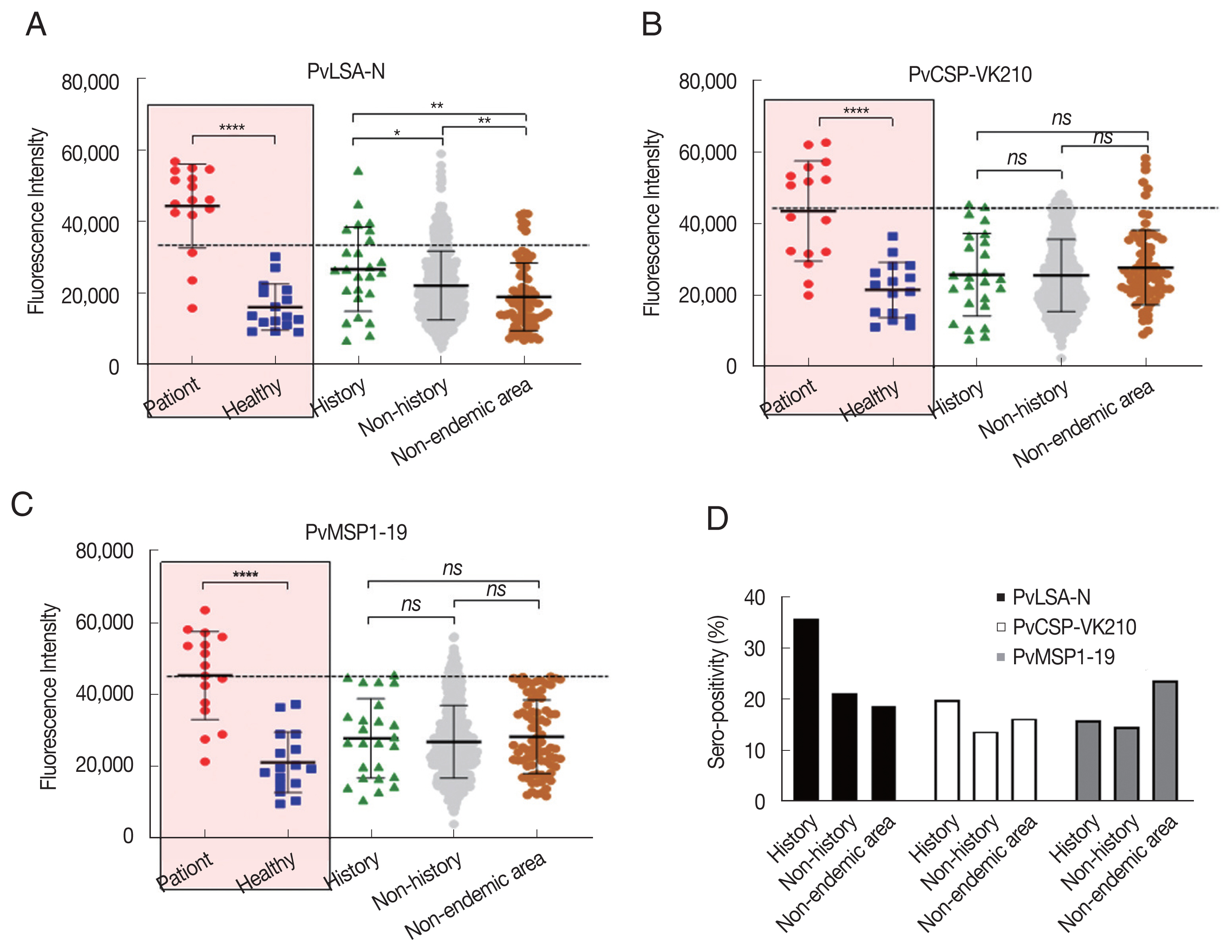
Fig. 1B). Seropositivity among the cases with a history was compared with those without a history and samples from nonendemic areas. Interestingly, there was a significant difference between history and nonhistory from endemic areas as well as the nonendemic area (Haman-gun) for PvLSA-N, but this was not observed for the other antigens (
Fig. 4A–C). In addition, seropositivity to PvLSA-N among history samples was much higher than that of nonhistory and nonendemic area samples (
Fig. 4D), suggesting that PvLSA-N is a protein marker to detect the specific antibody remaining in people who have been infected with the malaria parasite.
DISCUSSION
Elimination and control of the malaria parasite from endemic areas are dependent on the diagnosis of patients with or without symptoms. In particular, as asymptomatic malaria patients play an important role as carriers of the parasite, it identifying them by effective methods is key. However, the greatest obstacle to detecting and treating asymptomatic patients is that they do not need to visit the hospital or local health center [
13,
14]. Thus, blood samples of residents from 3 vivax malaria-endemic areas were collected, and the collected samples were examined by microscopy, RDT, and nested PCR methods to detect asymptomatic carriers in the endemic areas of Korea.
Microscopic examination and RDT are recommended for the detection of malaria infection at field sites. However, the sensitivity of the above methods for detecting low parasitemia is relatively lower than that of molecular diagnostic methods such as PCR-based techniques, and expert specialists are required for microscopic examination to identify and distinguish malaria species; moreover, the technique is time-consuming and can lead to misdiagnosis. In contrast, various PCR-based methods are currently well developed, and these techniques can detect a low level of the parasite that is not detected by blood film and/or RDT methods. Thus, 3 diagnostic methods were used in the present study to identify patients among the residents of malaria-endemic areas [
15,
16].
A total of 777 blood samples were collected from 3 endemic areas, Gimpo-si, Paju-si, and Yeoncheon-gun, which are located in the northwestern region bordering the demilitarized zone; a large number of clinical malaria cases have been reported in the previous 3 years compared with other areas. All collected samples were checked by microscopic examination, RDT, and nested PCR. However, no positive samples were detected, even though the amplified 18S rRNA gene of P. vivax parasites was observed as a single band in the positive control data not shown; thus, that there was no infection among the collected samples. There were 86 clinical malaria cases reported in Paju in 2019, including over 450,000 people and indicating that only 0.03% of people were infected with malaria in this city. Hence, the total number of tested samples from 3 endemic areas was not sufficient for determining asymptomatic patients and/or long-term relapse cases in this study, suggesting that mass-screening research must be scaled up to a larger population with continuous monitoring in these areas to detect malaria patients at the community level.
Malaria serological analysis of antibodies is crucial for estimating the local transmission of malaria in endemic areas, and various antigens have been used for serological and immune response analysis and antibody longevity [
17–
20]. In this study, PvMSP1-19, PvCSP-VK210, and PvLSA-N were used as antigens for serological analysis. MSP1-19 is a relatively conserved amino acid sequence fragment, and
P. falciparum can induce antibodies that prevent infection by blood-stage parasites as well as malaria-related febrile illness [
21]. Furthermore, the anti-MSP1-19 antibody is known to have protective activity against symptomatic malaria infection, and MSP1-19 is widely used as a serological marker. Moreover, there is a study in which antibodies were detected longer than 9 months after malaria infection, indicating that this antigen can be used to track the local transmission of malaria under a low-transmission setting [
22]. CSP is a sporogony-stage protein and is located on the surface membrane of sporozoites. Of the CSP fragments, the central repeat region contains multiple copies of the B-cell epitope; thus, it is highly immunogenic and can induce a protective response to sporozoite infection [
23]. However, the life-span of antibodies to CSP is just within 27 days, indicating that it is difficult to use this antigen to track historical malaria infection. In this study, there was no significant difference between the samples collected from those with and without a history of infection when using PvMSP1-19 and PvCSP-VK210.
In contrast, PvLSA-N could distinguish the 2 groups. According to this high level of positivity for PvLSA antigen in the present study (
Fig. 4A, D), it is possible to use PvLSA as a serological marker to track past infection of vivax malaria in individuals as well as the community. Nonetheless, the biological and functional activities of PvLSA in the erythrocytes and pre-erythrocytes of vivax malaria parasites have not yet been described. Thus, it may also be necessary to define PvLSA for diagnostic applications in vivax endemic areas.
In our surveyed areas, we found that one patient developed febrile symptoms approximately one month after our sample collection. However, in our tests, parasites were not detected by 3 methods, including the molecular detection method, and a high antibody titer from serological assays was not found during the nonmalaria endemic season before the emergence of mosquitoes for malaria transmission. It is possible to infect by biting new mosquitoes or developing parasites from relapsed parasites after our examination. Thus, epidemiological surveillance should be sustained in endemic areas, and alternative ways to detect long- and short-relapse cases need to be found.
In summary, a total of 777 blood samples were collected from residents in 3 vivax malaria-endemic areas of Korea and examined by microscopy, RDTs, and PCR to detect malaria. However, there were no positive malaria cases, which indicates that a more substantial number of samples are required for further epidemical surveillance. Based on serological analysis of samples using 3 P. vivax antigens, we identified that PvLSA-N can be used as a serological marker to detect pre-exposure to malaria infection. In the future, P. vivax serological markers should be identified in well-characterized field samples.
Notes
-
CONFLICT OF INTEREST
The authors declare no conflict of interest related to this study.
ACKNOWLEDGMENT
This work was supported by the Korea Centers for Disease Control and Prevention (2019E440100), Korea.
Fig. 1Sample collection from vivax malaria endemic and non-endemic areas in Korea. (A) Map shows vivax malaria-endemic areas (enlarged) and non-endemic area, Haman-gun. (B) Gender and past history of residents enrolled in the endemic areas.
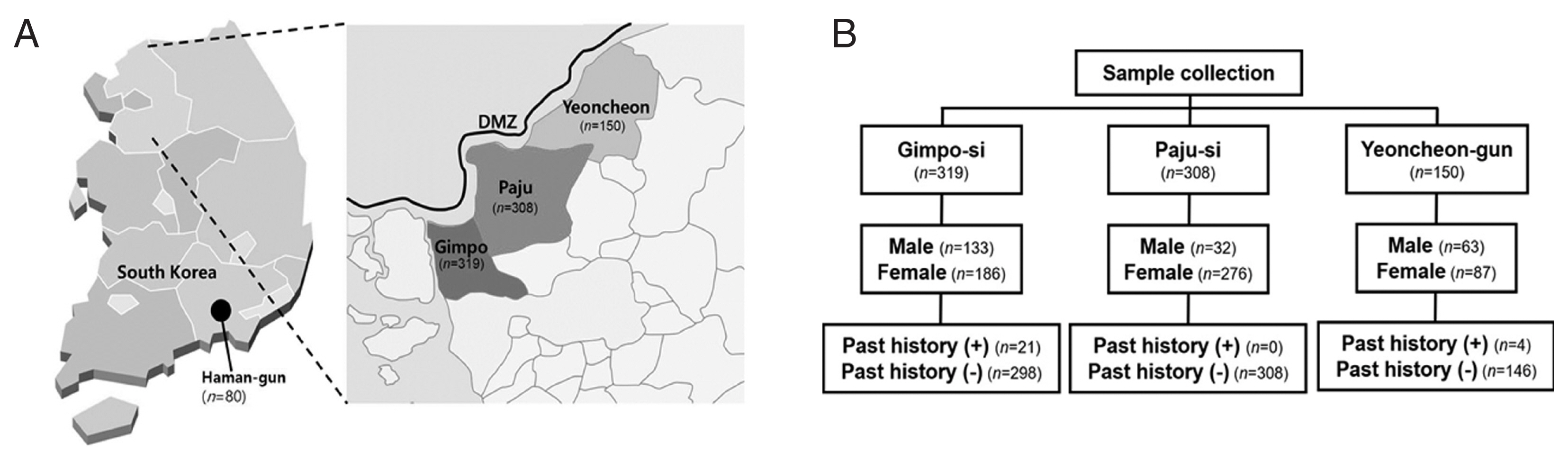
Fig. 2Recombinant protein domains selected for Plasmodium vivax recombinant proteins. (A) Schematic structures of Plasmodium vivax merozoite surface protein 1 (PvMSP1) and P. vivax liver-stage antigen (PvLSA). (B) Full-length, including a combination of central repeat region sequence of PvCSP. (C) A western-blot of 3 recombinant proteins in wheat-germ cell-free expression system.
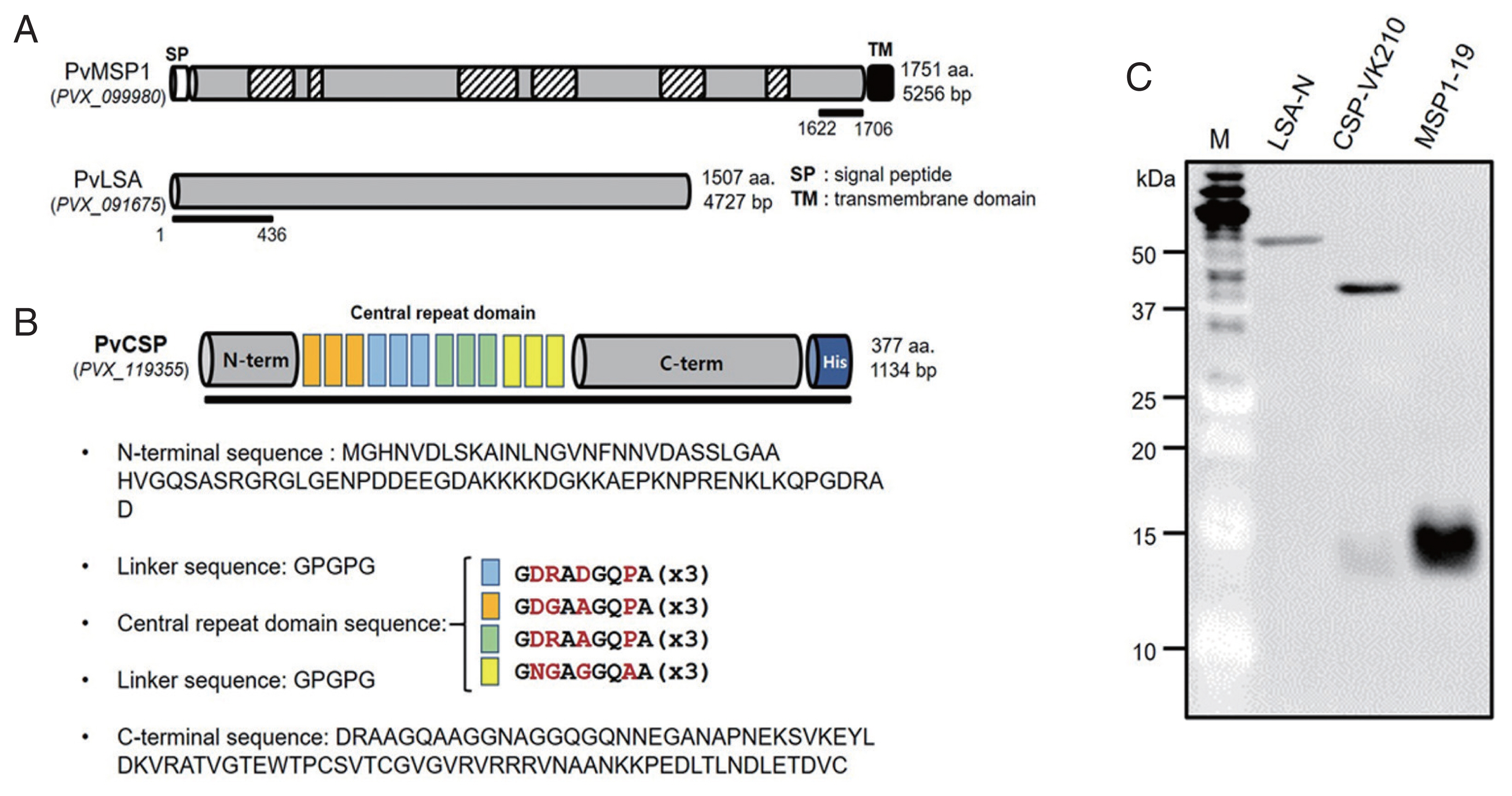
Fig. 3Fluorescence intensity of sera to PvLSA-N (A), PvCSP-VK210 (B), and PvMSP1-19 (C), collected from 3 study areas, Gimpo-si, Paju-si, and Yeoncheon-gun and a non-endemic area, Haman-gun.
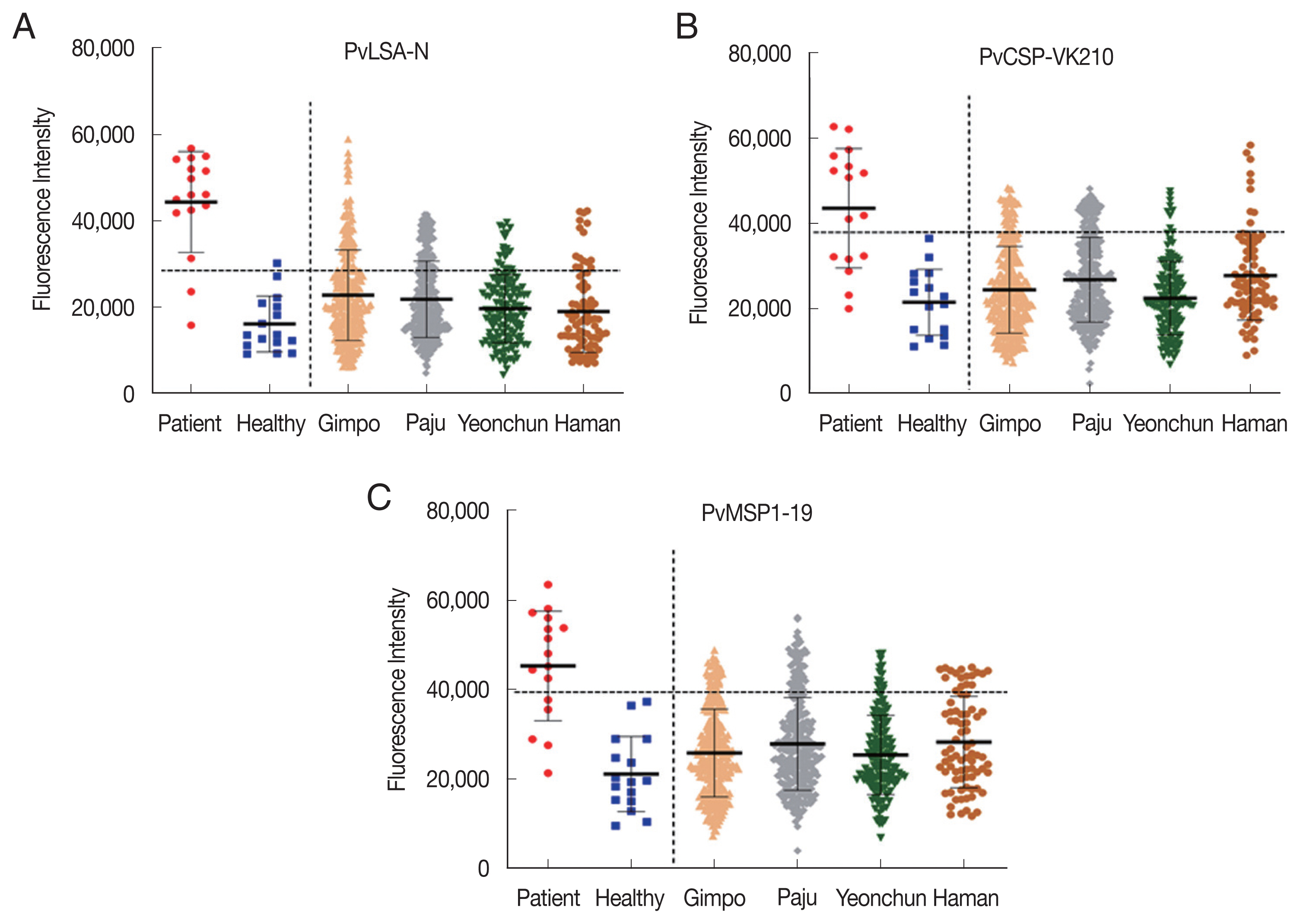
Fig. 4Seroprevalence of 3 Plasmodium vivax recombinant antigens. PvLSA-N (A), PvCSP-VK210 (B), and PvMSP1-19 (C) referred to malaria infection history in the endemic areas and non-endemic area. (D) Seropositivity comparison of antigens between history and non-history in endemic and non-endemic areas. ****, P<0.0001; ***, P<0.01; **, P<0.05; ns, not significance.
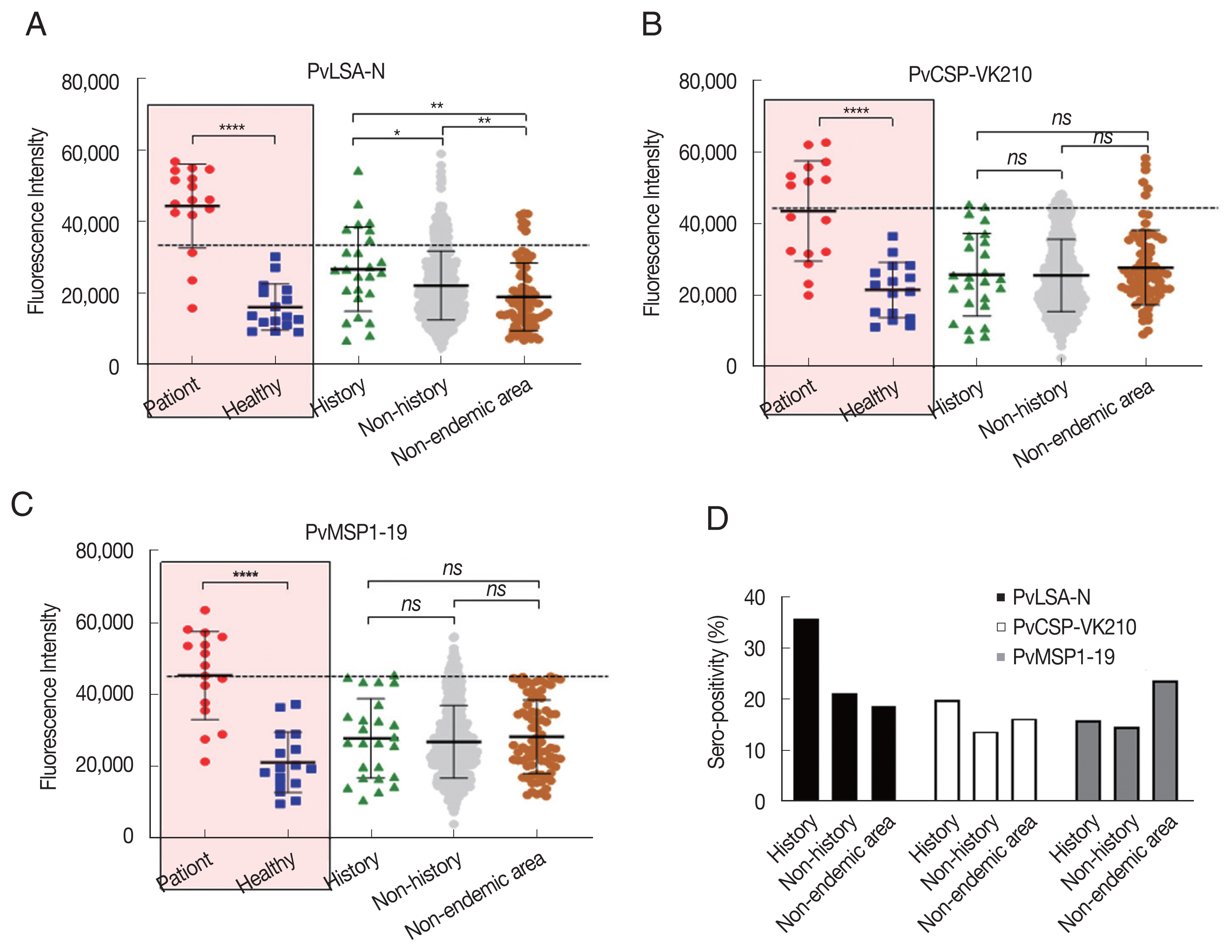
Table 1Reactivity of three Plasmodium vivax recombinant antigens toward control sera of vivax patients and healthy individuals
Table 1
|
Antigen |
Sera of vivax patients |
MFI |
SD |
Sera of healthy individuals |
MFI |
SD |
|
|
|
Positive |
Negative |
Sensitivity (%) |
Positive |
Negative |
Specificity (%) |
|
PvLSA-N |
14 |
2 |
87.5 |
44,356 |
11,673 |
1 |
15 |
93.8 |
16,103 |
6,454 |
|
|
PvCSP-VK210 |
10 |
6 |
62.5 |
43,579 |
14,021 |
0 |
16 |
100 |
21,476 |
7,736 |
|
|
PvMSP1-19 |
11 |
5 |
68.7 |
45,306 |
12,265 |
0 |
16 |
100 |
21,106 |
8,404 |
Table 2Positivity of sera from residents in vivax endemic areas against three recombinant Plasmodium vivax antigens
Table 2
|
Antigen |
Region |
Total |
Positive (%) |
MFI |
SD |
|
PvLSA-N |
Gimpo-si |
319 |
23.3 |
22,781 |
10,488 |
|
Paju-si |
308 |
22.7 |
21,818 |
8,866 |
|
Yeoncheon-gun |
150 |
10.7 |
19,646 |
7,883 |
|
Subtotala
|
777 |
20.2±6.8 |
|
|
|
Haman |
80 |
18.8 |
18,972 |
9,477 |
|
|
PvCSP-VK210 |
Gimpo-si |
319 |
15.4 |
24,392 |
10,193 |
|
Paju-si |
308 |
18.8 |
26,766 |
9,952 |
|
Yeoncheon-gun |
150 |
6.7 |
22,421 |
8,636 |
|
Subtotal |
777 |
15.1±6.3 |
|
|
|
Haman |
80 |
16.3 |
27,763 |
10,468 |
|
|
PvMSP1-19 |
Gimpo-si |
319 |
14.7 |
25,822 |
9,818 |
|
Paju-si |
308 |
19.5 |
27,845 |
10,385 |
|
Yeoncheon-gun |
150 |
10.0 |
25,391 |
8,877 |
|
Subtotal |
777 |
15.7±4.7 |
|
|
|
Haman |
80 |
23.8 |
36,897 |
14,068 |
References
- 1. Mendis K, Sina BJ, Marchesini P, Carter R. The neglected burden of Plasmodium vivax malaria. Am J Trop Med Hyg 2001;64:97-106. https://doi.org/10.4269/ajtmh.2001.64.97
- 2. Chai JY. Re-emerging Plasmodium vivax malaria in the Republic of Korea. Korean J Parasitol 1999;37:129-143. https://doi.org/10.3347/kjp.1999.37.3.129
- 3. Sturrock HJ, Hsiang MS, Cohen JM, Smith DL, Greenhouse B, Bousema T, Gosling RD. Targeting asymptomatic malaria infections: active surveillance in control and elimination. PLoS Med 2013;10:e1001467. https://doi.org/10.1371/journal.pmed.1001467
- 4. White NJ. Determinants of relapse periodicity in Plasmodium vivax malaria. Malar J 2011;10:297. https://doi.org/10.1186/1475-2875-10-297
- 5. Warhurst DC, Williams JE. ACP Broadsheet no 148. July 1996. Laboratory diagnosis of malaria. J Clin Pathol 1996;49:533-538. https://doi.org/10.1136/jcp.49.7.533
- 6. Kain KC, Harrington MA, Tennyson S, Keystone JS. Imported malaria: prospective analysis of problems in diagnosis and management. Clin Infect Dis 1998;27:142-149. https://doi.org/10.1086/514616
- 7. Komaki-Yasuda K, Vincent JP, Nakatsu M, Kato Y, Ohmagari N, Kano S. A novel PCR-based system for the detection of four species of human malaria parasites and Plasmodium knowlesi. PLoS One 2018;13:e0191886. https://doi.org/10.1371/journal.pone.0191886
- 8. Nyunt MH, Soe TN, Shein T, Zaw NN, Han SS, Muh F, Lee SK, Han JH, Park JH, Ha KS, Park WS, Hong SH, Kyaw MP, Han ET. Estimation on local transmission of malaria by serological approach under low transmission setting in Myanmar. Malar J 2018;17:6. https://doi.org/10.1186/s12936-017-2170-8
- 9. Nyunt MH, Kyaw MP, Thant KZ, Shein T, Han SS, Zaw NN, Han JH, Lee SK, Muh F, Kim JY, Cho SH, Lee SE, Yang EJ, Chang CL, Han ET. Effective high-throughput blood pooling strategy before DNA extraction for detection of malaria in low-transmission settings. Korean J Parasitol 2016;54:253-259. https://doi.org/10.3347/kjp.2016.54.3.253
- 10. Ndao M, Bandyayera E, Kokoskin E, Gyorkos TW, MacLean JD, Ward BJ. Comparison of blood smear, antigen detection, and nested-PCR methods for screening refugees from regions where malaria is endemic after a malaria outbreak in Quebec, Canada. J Clin Microbiol 2004;42:2694-2700. https://doi.org/10.1128/JCM.42.6.2694-2700.2004
- 11. Tsuboi T, Takeo S, Iriko H, Jin L, Tsuchimochi M, Matsuda S, Han ET, Otsuki H, Kaneko O, Sattabongkot J, Udomsangpetch R, Sawasaki T, Torii M, Endo Y. Wheat germ cell-free system-based production of malaria proteins for discovery of novel vaccine candidates. Infect Immun 2008;76:1702-1708. https://doi.org/10.1128/IAI.01539-07
- 12. Chen JH, Jung JW, Wang Y, Ha KS, Lu F, Lim CS, Takeo S, Tsuboi T, Han ET. Immunoproteomics profiling of blood stage Plasmodium vivax infection by high-throughput screening assays. J Proteome Res 2010;9:6479-6489. https://doi.org/10.1021/pr100705g
- 13. Hassanpour G, Mohebali M, Zeraati H, Raeisi A, Keshavarz H. Asymptomatic malaria and its challenges in the malaria elimination program in Iran: a systematic review. J Arthropod Borne Dis 2017;11:172-181.
- 14. Zhao Y, Zeng J, Liu Q, He Y, Zhang J, Yang Z, Fan Q, Wang Q, Cui L, Cao Y. Risk factors for asymptomatic malaria infections from seasonal cross-sectional surveys along the China-Myanmar border. Malar J 2018;17:247. https://doi.org/10.1186/s12936-018-2398-y
- 15. Wang B, Han SS, Cho C, Han JH, Cheng Y, Lee SK, Galappaththy GN, Thimasarn K, Soe MT, Oo HW, Kyaw MP, Han ET. Comparison of microscopy, nested-PCR, and Real-Time-PCR assays using high-throughput screening of pooled samples for diagnosis of malaria in asymptomatic carriers from areas of endemicity in Myanmar. J Clin Microbiol 2014;52:1838-1845. https://doi.org/10.1128/JCM.03615-13
- 16. Yin J, Li M, Yan H, Zhou S. Considerations on PCR-based methods for malaria diagnosis in China malaria diagnosis reference laboratory network. Biosci Trends 2018;12:510-514. https://doi.org/10.5582/bst.2018.01198
- 17. Cassiano GC, Furini AA, Capobianco MP, Storti-Melo LM, Almeida ME, Barbosa DR, Póvoa MM, Nogueira PA, Machado RL. Immunogenetic markers associated with a naturally acquired humoral immune response against an N-terminal antigen of Plasmodium vivax merozoite surface protein 1 (PvMSP-1). Malar J 2016;15:306. https://doi.org/10.1186/s12936-016-1350-2
- 18. Baum E, Sattabongkot J, Sirichaisinthop J, Kiattibutr K, Jain A, Taghavian O, Lee MC, Huw Davies D, Cui L, Felgner PL, Yan G. Common asymptomatic and submicroscopic malaria infections in Western Thailand revealed in longitudinal molecular and serological studies: a challenge to malaria elimination. Malar J 2016;15:333. https://doi.org/10.1186/s12936-016-1393-4
- 19. Chen E, Salinas ND, Huang Y, Ntumngia F, Plasencia MD, Gross ML, Adams JH, Tolia NH. Broadly neutralizing epitopes in the Plasmodium vivax vaccine candidate Duffy Binding Protein. Proc Natl Acad Sci USA 2016;113:6277-6282. https://doi.org/10.1073/pnas.1600488113
- 20. Drakeley CJ, Corran PH, Coleman PG, Tongren JE, McDonald SL, Carneiro I, Malima R, Lusingu J, Manjurano A, Nkya WM, Lemnge MM, Cox J, Reyburn H, Riley EM. Estimating medium- and long-term trends in malaria transmission by using serological markers of malaria exposure. Proc Natl Acad Sci USA 2005;102:5108-5113. https://doi.org/10.1073/pnas.0408725102
- 21. Branch OH, Udhayakumar V, Hightower AW, Oloo AJ, Hawley WA, Nahlen BL, Bloland PB, Kaslow DC, Lal AA. A longitudinal investigation of IgG and IgM antibody responses to the merozoite surface protein-1 19-kiloDalton domain of Plasmodium falciparum in pregnant women and infants: associations with febrile illness, parasitemia, and anemia. Am J Trop Med Hyg 1998;58:211-219. https://doi.org/10.4269/ajtmh.1998.58.211
- 22. Teo A, Feng G, Brown GV, Beeson JG, Rogerson SJ. Functional antibodies and protection against blood-stage malaria. Trends Parasitol 2016;32:887-898. https://doi.org/10.1016/j.pt.2016.07.003
- 23. Arnot DE, Barnwell JW, Tam JP, Nussenzweig V, Nussenzweig RS, Enea V. Circumsporozoite protein of Plasmodium vivax: gene cloning and characterization of the immunodominant epitope. Science 1985;230:815-818. https://doi.org/10.1126/science.2414847

 , Sang-Eun Lee3, Hyun-Il Shin3, Shin Hyeong Cho3, Won Sun Park4, Feng Lu2, Eun-Taek Han1,*
, Sang-Eun Lee3, Hyun-Il Shin3, Shin Hyeong Cho3, Won Sun Park4, Feng Lu2, Eun-Taek Han1,*