Abstract
Astrocytes are the most abundant cells in the central nervous system that play roles in maintaining the blood-brain-barrier and in neural injury, including cerebral malaria, a severe complication of Plasmodium falciparum infection. Prostaglandin (PG) D2 is abundantly produced in the brain and regulates the sleep response. Moreover, PGD2 is a potential factor derived from P. falciparum within erythrocytes. Heme oxygenase-1 (HO-1) is catalyzing enzyme in heme breakdown process to release iron, carbon monoxide, and biliverdin/bilirubin, and may influence iron supply to the P. falciparum parasites. Here, we showed that treatment of a human astrocyte cell line, CCF-STTG1, with PGD2 significantly increased the expression levels of HO-1 mRNA by RT-PCR. Western blot analysis showed that PGD2 treatment increased the level of HO-1 protein, in a dose- and time-dependent manner. Thus, PGD2 may be involved in the pathogenesis of cerebral malaria by inducing HO-1 expression in malaria patients.
-
Key words: Plasmodium falciparum, astrocyte, heme oxygenase, iron, cerebral malaria, prostaglandin D2
INTRODUCTION
Astrocytes are the most numerous cell types in the central nervous system (CNS). They provide structural, trophic, and metabolic support to neurons and modulate synaptic activity [
1]. Astrocytes were used in various studies involving in neurological diseases [
2-
5]. These cells have the most direct interaction with vasculature, and therefore their endfeet are in contact with cerebral endothelial cells (CECs) [
6,
7]. In vitro and in vivo studies support the role of astrocytes in controlling blood-brain-barrier (BBB) maintenance and regulation through their interaction with CECs [
8-
12]. The neurological dysfunction caused by breakdown of BBB thereby allowing compounds, such as histamine and reactive oxygen species (ROS) to enter the brain, may lead to the pathological changes in the brain [
13].
Malaria is a worldwide protozoan infection [
14], and most malignant malaria is caused by
Plasmodium falciparum. Cerebral malaria is one of the most severe complications of
P. falciparum infection [
15,
16]. The parasite does not enter the brain parenchyma, but staying in the intravascular circulation, which is responsible for changes in the BBB. Cerebral malaria is characterized by a sequestration of parasitized red blood cells in the brain microvasculature. The sequestration of these parasitized erythrocytes could result in activation of cerebral endothelial cells [
17]. Perivascular macrophages (perivascular cells) are amongst the first cells to encounter proteins leaking across a disrupted BBB, and as a consequence of activation or phagocytosis, they may secrete a wide range of pro-inflammatory and neuroactive mediators which could influence local neuronal functions. It has been reported that, during cerebral malaria, the cerebral parenchyma is not largely affected, and local events occur within and around the cerebral microvasculature [
18]. The molecular basis underlying cerebral malaria, nevertheless, remains unclear.
Recently, heme oxygenase (HO) has been proposed as one of the factors that play significant roles in the pathogenesis of falciparum malaria complication [
19,
20]. It is a microsomal enzyme which exists in 2 isoforms, i.e., HO-1 and HO-2. Heme oxygenases are rate-limiting enzymes in heme catabolism to generate biliverdin IXα/bilirubin IXα, carbon monoxide, and ferrous iron [
20-
22]. The expression levels of HO-1 is inducible or repressible, depending on cell types or cellular microenvironments, but expression levels of HO-2 are fairly constant [
20,
22]. The main regulation of heme catabolism is, therefore, determined by the balance between induction and repression of HO-1.
HO-1 levels in the brain were studied in Alzheimer's (AD), Parkinson's (PD), and infectious diseases, including malaria [
23,
24]. It has been proposed that the mechanisms for the fine-tuning of HO-1 expression may involve the polymorphic (GT)
n sequence (n = 15-40) located in the human HO-1 gene promoter [
23,
25]. We have recently reported that the short (GT)
n repeats (n < 28) in the HO-1 gene promoter are associated with higher incidence of cerebral malaria in the Karen ethnic minority group who live near the border between Myanmar and Thailand [
26].
Prostaglandins (PGs) originate from the degradation of membrane arachidonic acid by cyclooxygenases (COX-1 and COX-2). The prostaglandin actions in the nervous system have been suggested to play a significant role in neurodegenerative disorders [
27] and to be involved in various symptoms associated with parasitic diseases [
28]. PGD
2 is a major prostanoid produced in the brain and is involved in the regulation of sleep and pain responses [
29,
30]. Lipocalin-type PGD synthase (L-PGDS) catalyzes the isomerization of PGH
2 to produce PGD
2, and is mainly responsible for production of PGD
2 in the brain [
30,
31]. PGD
2 exerts its actions through the G protein coupled receptors DP1 and chemoattractant-homologous receptor expressed on Th2 cells (CRTH2, also known as DP2) which are expressed in various cell types [
32-
34].
In 1998, Kubata et al. [
28] have shown that the cell homogenates of
P. falciparum contain the activity that produces PGD
2 and PGE
2 after incubation with arachidonic acid. Notably, PGD
2 was shown to be the predominant accumulate in the culture medium of the parasitized erythrocytes after treatment with arachidonic acid [
28]. Moreover, PGD
2 was maintained at a higher level in the serum of falciparum malaria patients than the control serum [
35]. We previously reported that PGD
2 and 15d-PGJ
2, a minor species of PGD
2 metabolites, were found to increase HO-1 gene promoter, mRNA levels, and protein levels in retinal pigment epithelial cells that may associate with the pathogenesis of malarial retinopathy in malaria patients with cerebral complication [
36,
37].
In the present study, we provide additional data to support the link between HO-1 and pathogenesis of cerebral malaria through the induction by PGD2 in human astrocytes.
MATERIALS AND METHODS
Cell culture
The human astrocyte cell line, CCF-STTG1 was purchased from American Type Culture Collection (CRL-1718, ATCC, Virginia, USA). CCF-STTG1 cells were cultured in RPMI 1640 medium, supplemented with 10% fetal bovine serum (FBS), 2 mM L-glutamine and antibiotics (100 U/ml penicillin and 0.1 mg/ml streptomycin). To examine the effects of PGD2 and 15d-PGJ2 (Cayman Chemicals, Michigan, USA) on the expression levels of HO-1 protein and mRNA, CCF-STTG1 cells were grown to 70-80% confluence before they were incubated with vehicle (ethanol), PGD2 or 15d-PGJ2.
Western blot analysis
Cells were lysed in the buffer containing 20 mM Hepes (pH 7.9), 400 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1.5 mM MgCl
2, 1 mM DTT, 0.1% protease inhibitor cocktail (Sigma-Aldrich, St. Louis, Missouri, USA) and 0.5% Nonidet P-40 (Fluka, Buchs, Switzerland) [
36]. The resultant lysates were centrifuged at 15,000 g for 5 min at 4℃ after incubation for 15 min, and then supernatants were collected. Lysate supernatant proteins (30 µg per lane) were fractionated by SDS-PAGE and blotted to a nitrocellulose membrane (Hybond ECL™, Amersham, New Jersey, USA) in buffer containing 20% methanol, 48 mM Tris, 39 mM glycine, and 0.037% SDS. The membranes (Western blots) were treated 1-2 hr in Tris-buffered saline (TBS), containing 5% non-fat dried milk, and were washed 3 times each for 10 min in TBS with 0.1% Tween 20 (TBS-T) at room temperature. The proteins were probed with HO-1 (SPA-895, StressGen Biotechnologies) and HO-2 (SPA-897, StressGen Biotechnologies, Michigan, USA) antibodies, and β-actin antibody (Sigma) at a dilution of 1 : 1,000 for 1 hr at room temperature. For detection of HO-1 and HO-2 proteins, the blots were incubated with alkaline phosphatase (AP)-goat anti-rabbit antibodies (ZyMax™, Invitrogen, California, USA), and β-actin protein was incubated with AP-goat anti-mouse antibodies (ZyMax™, Invitrogen). The color was developed by incubating the blot in the developing solution, BCIP/NBT (AMRESCO, Ohio, USA).
Total RNA was extracted from treated CCF-STTG1 cells using RNeasy® Mini Kit (Qiagen, Hilden, Germany). Total RNA was transcribed to cDNA using Omniscript® RT Kit (Qiagen). Then, HO-1 cDNA levels were determined by RT-PCR. Primers were designed according to the published cDNA sequences for human HO-1 [
38] and HO-2 [
39]. The forward 5'-CAGGCAGAGAATGCTGAG-3'and the reverse 5'-GCTTCACATAGCGCTGCA-3' primers for HO-1 were used to amplify a 271-bp fragment, and the forward 5'-ATGTCAGCGGAAGTGGAA-3'and the reverse 5'-GGGAGTTTCAGTGCTCGC-3'primers for HO-2 were used to amplify a 533-bp fragment. The house-keeping gene, glyceraldehyde-3-phosphate dehydrogenase, was also amplified using the following primers 5'-TGAAGGTCGGAGTCAACGGATTTG-3'and 5'-GCGCCAGTAGAGGCAGGGATGATG-3', yielding a 628-bp product [
40].
The cDNA template were amplified with an initial hold for 5 min at 95℃, followed by 40 cycles of denaturation at 95℃ for 15 sec, and annealing/extension at 60℃ for 60 sec in iCycler iQ real time PCR (Bio-Rad, California, USA). Relative HO-1 mRNA and HO-2 mRNA expression were obtained by dividing the intensity value for each sample with 0-hr untreated control culture cells, which reflected basal expression level (normalized to 1.0) by using glyceraldehyde-3-phosphate dehydrogenase for standardization. Data were analyzed in duplicate.
RESULTS
Effects of PGD2 on the expression of HO-1 and HO-2 proteins in CCF-STTG1 cells
We performed the time-course studies of the effects of PGD
2 on the expression of HO-1 and HO-2 proteins in CCF-STTG1 at 3, 6, 12, and 24 hr, and dose-response studies of PGD
2 at a final concentration of 0.5, 1, 5, or 10 µM in CCF-STTG1 cells. Western blot analysis revealed that the expression of HO-1 protein was induced in a dose-dependent manner by PGD
2 at a final concentration of 5 and 10 µM after a 24-hr treatment (
Fig. 1A). The apparent induction of HO-1 protein was detected at 12 and 24 hr with the concentration of PGD
2 5 and 10 µM. In contrast, the expression level of HO-2 protein was unchanged or did not increase after a 24-hr treatment with the highest concentration of PGD
2 (10 µM). The expression of β-actin, an internal control was unchanged. These results indicate that PGD
2 induces the expression of HO-1 protein in CCF-STTG1 cells.
It has been well established that 15d-PGJ
2 increases transcription of the HO-1 gene [
36,
41,
42]. We, therefore, performed a similar experiment with 15d-PGJ
2 for comparison. As expected, 15d-PGJ
2 induced the expression of HO-1 protein in a dose-dependent and time-dependent manner in CCF-STTG1 cells (
Fig. 1B). The apparent induction of HO-1 was detected after a 6-hr treatment with 15d-PGJ
2 (5 and 10 µM) (
Fig. 1B). The onset of the HO-1 induction with 15d-PGJ
2 was earlier than that with PGD
2. HO-2 as well as β-actin protein levels remained unchanged after treatment with 15d-PGJ
2.
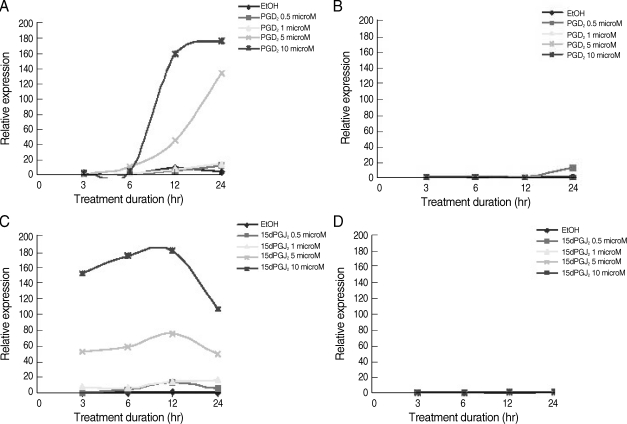
We next performed the time-course and dose-response effects of PGD
2 and 15d-PGJ
2 on the expression of HO-1 mRNA in C CF-STTG1 cells using RT-PCR. The expression of HO-1 mRNA was induced by PGD
2 or 15d-PGJ
2 in a dose-dependent manner (
Fig. 2). The expression levels of HO-1 mRNA were increased after 6 hr of the treatment with PGD
2 of 5 µM or 10 µM, and continuously increased at maximum levels after 12 hr (
Fig. 2A). For 15d-PGJ
2, the expression levels of HO-1 mRNA were detected at 3 hr of treatment (
Fig. 2C). The maximum induction of HO-1 mRNA was achieved after 6 hr of treatment, but it was decreased at 24 hr (
Fig. 2C). In contrast, HO-2 mRNA levels remained unchanged in both of induction (
Fig. 2B, D). The induction profiles of HO-1 mRNA were in good agreement with those of HO-1 protein (
Fig. 1).
DISCUSSION
The astrocytes are known as cells surrounding cerebral endothelial cells [
43] that influence the BBB function [
12,
44,
45] by releasing cytokines and other soluble factors which may induce adverse responses in surrounding neuronal tissues [
46,
47]. In addition, astrocytes have the property to synthesize prostaglandins [
48,
49].
The endothelial cells also have a reciprocal inductive influence on astrocytes [
50,
51]. Therefore, the astrocyte-endothelial interaction may be involved in the pathogenesis of encephalopathy caused by
P. falciparum. Most of the antimalarial agents used in the treatment of cerebral malaria (quinine, artemether, artesunate, and mefloquine) have been reported to cause neurotoxicity [
52-
58]. The integrity of the BBB, including the functionality of the cerebral efflux proteins, e.g., p-glycoproten (P-gp), multidrug resistant protein-1 (MRP1), or breast cancer resistant protein (BCRP) are therefore important factors in cerebral malaria to control the cerebral transport and the neurotoxicity of these drugs [
59-
62]. Nevertheless, little information is available regarding the effects of alteration of BBB integrity in cerebral malaria as well as interactions of antimalarial agents on antimalarial toxicity [
14,
17,
63,
64].
The present results showing that, the treatment of astrocyzte cells CCF-STTG1 with exogenous PGD
2 consistently induced HO-1 expression, is in agreement with our recent report in retinal pigment epithelial cells [
36]. Endogenously produced PGD
2 by astrocyte cells may contribute to the appropriate expression of HO-1 in the brain. The induction profile of HO-1 expression with PGD
2 is essentially similar to that with 15d-PGJ
2, although the onset of the induction by PGD
2 is later than by 15d-PGJ
2. It remains to be explored whether PGD
2 itself or a specific PGD
2 metabolite, other than 15d-PGJ
2, is responsible for the induction of HO-1 expression. Apart from astrocyte cells themselves,
falciparum parasites may also release PGD
2 [
28,
48] that may hasten the expression of HO-1. This consequence additively results in the increase in HO-1 activity to catalyze heme to end products, especially, iron. Malaria growth and proliferation depend on iron supply from host cells, such as endothelial cells or neuron cells near the sequestration site. Therefore, PGD
2 might enhance the growth of parasites by modulating iron availability from the host. The stimulation of HO-1 by PGD
2 and the metabolite 15d-PGJ
2 observed in this study is critical since excessive heme degradation may result in toxic levels of iron similar to that by carbon monoxide, and bilirubin/biliverdin.
Detailed studies are underway in order to provide evidence to definitely conclude on the prostaglandin-mediated stimulatory effects on HO-1 and its role in malaria pathogenesis. Preliminary studies showed significant findings. These include investigation of (i) the mechanism of PGD2-mediated induction of HO-1 expression in pigment epithelial cell lines (ARPE-19 and D407 RPE) in vitro, (ii) the production of PGD2 in P. falciparum culture in vitro, (iii) the growth inhibitory effects of the antagonists of PGD2 receptors (DP1 and DP2 antagonists) on HUVEC cells when cocultured with P. falciparum in vitro, and (iv) the association between PGD2 levels and malaria pathogenesis in patients. The knowledge on the link of the polymorphism of GT repeat of HO-1 gene promoter in modulating the enzyme activity of HO-1 and susceptibility to severe malaria may be exploited for further development of new drugs acting as inhibitors of HO-1 to prevent the progression to severe cerebral malaria.
ACKNOWLEDGEMENTS
This work was supported by Thammasat University and The Commission on Higher Education, Ministry of Education of Thailand.
References
- 1. Chen Y, Swanson RA. Astrocytes and brain injury. J Cereb Blood Flow Metab 2003;23:137-149.
- 2. Diedrich JF, Minnigan H, Carp RI, Whitaker JN, Race R, Frey W 2nd, Haase AT. Neuropathological changes in scrapie and Alzheimer's disease are associated with increased expression of apolipoprotein E and cathepsin D in astrocytes. J Virol 1991;65:4759-4768.
- 3. Murayama S, Inoue K, Kawakami H, Bouldin TW, Suzuki K. A unique pattern of astrocytosis in the primary motor area in amyotrophic lateral sclerosis. Acta Neuropathol 1991;82:456-461.
- 4. Aloisi F, Borsellino G, Caré A, Testa U, Gallo P, Russo G, Peschle C, Levi G. Cytokine regulation of astrocyte function: in-vitro studies using cells from the human brain. Int J Dev Neurosci 1995;13:265-274.
- 5. Mattson MP, Chan SL. Neuronal and glial calcium signaling in Alzheimer's disease. Cell Calcium 2003;34:385-397.
- 6. Abbott NJ. Astrocyte-endothelial interactions and blood-brain barrier permeability. J Anat 2002;200:629-638.
- 7. Ballabh P, Braun A, Nedergaard M. The blood-brain barrier: an overview: structure, regulation, and clinical implications. Neurobiol Dis 2004;16:1-13.
- 8. Maxwell K, Berliner JA, Cancilla PA. Induction of gamma-glutamyl transpeptidase in cultured cerebral endothelial cells by a product released by astrocytes. Brain Res 1987;410:309-314.
- 9. Neuhaus J, Risau W, Wolburg H. Induction of blood-brain barrier characteristics in bovine brain endothelial cells by rat astroglial cells in transfilter coculture. Ann N Y Acad Sci 1991;633:578-580.
- 10. Hayashi Y, Nomura M, Yamagishi S, Harada S, Yamashita J, Yamamoto H. Induction of various blood-brain barrier properties in non-neural endothelial cells by close apposition to co-cultured astrocytes. Glia 1997;19:13-26.
- 11. Sobue K, Yamamoto N, Yoneda K, Hodgson ME, Yamashiro K, Tsuruoka N, Tsuda T, Katsuya H, Miura Y, Asai K, Kato T. Induction of blood-brain barrier properties in immortalized bovine brain endothelial cells by astrocytic factors. Neurosci Res 1999;35:155-164.
- 12. Janzer RC, Raff MC. Astrocytes induce blood-brain barrier properties in endothelial cells. Nature 1987;325:253-257.
- 13. Clark IA, Ilschner S, MacMicking JD, Cowden WB. TNF and Plasmodium berghei ANKA-induced cerebral malaria. Immunol Lett 1990;25:195-198.
- 14. Na-bangchang K, Congpoung K. Current malaria status and distribution of drug resistance in East and Southeast Asia with special focus to Thailand. Tohoku J Exp Med 2007;211:99-113.
- 15. Ma N, Madigan MC, Chan-Ling T, Hunt NH. Compromised blood-nerve barrier, astrogliosis, and myelin disruption in optic nerves during fatal murine cerebral malaria. Glia 1997;19:135-151.
- 16. Blanco YC, Farias AS, Goelnitz U, Lopes SC, Arrais-Silva WW, Carvalho BO, Amino R, Wunderlich G, Santos LM, Giorgio S, Costa FT. Hyperbaric oxygen prevents early death caused by experimental cerebral malaria. PLoS ONE 2008;3:e3126.
- 17. Brown H, Hien TT, Day N, Mai NT, Chuong LV, Chau TT, Loc PP, Phu NH, Bethell D, Farrar J, Gatter K, White N, Turner G. Evidence of blood-brain barrier dysfunction in human cerebral malaria. Neuropathol Appl Neurobiol 1999;25:331-340.
- 18. Brown HC, Chau TT, Mai NT, Day NP, Sinh DX, White NJ, Hien TT, Farrar J, Turner GD. Blood-brain barrier function in cerebral malaria and CNS infections in Vietnam. Neurology 2000;55:104-111.
- 19. Shibahara S, Kitamuro T, Takahashi K. Heme degradation and human disease: diversity is the soul of life. Antioxid Redox Signal 2002;4:593-602.
- 20. Shibahara S. The heme oxygenase dilemma in cellular homeostasis: new insights for the feedback regulation of heme catabolism. Tohoku J Exp Med 2003;200:167-186.
- 21. Shibahara S, Han F, Li B, Takeda K. Hypoxia and heme oxygenases: Oxygen sensing and regulation of expression. Antioxid Redox Signal 2007;9:2209-2225.
- 22. Furuyama K, Kaneko K, Vargas PD. Heme as a magnificent molecule with multiple missions: heme determines its own fate and governs cellular homeostasis. Tohoku J Exp Med 2007;213:1-16.
- 23. Kimpara T, Takeda A, Watanabe K, Itoyama Y, Ikawa S, Watanabe M, Arai H, Sasaki H, Higuchi S, Okita N, Takase S, Saito H, Takahashi K, Shibahara S. Microsatellite polymorphism in the human heme oxygenase-1 gene promoter and its application in association studies with Alzheimer and Parkinson disease. Hum Genet 1997;100:145-147.
- 24. Schluesener HJ, Kremsner PG, Meyermann R. Heme oxygenase-1 in lesions of human cerebral malaria. Acta Neuropathol 2001;101:65-68.
- 25. Yamada N, Yamaya M, Okinaga S, Nakayama K, Sekizawa K, Shibahara S, Sasaki H. Microsatellite polymorphism in the heme oxygenase-1 gene promoter is associated with susceptibility to emphysema. Am J Hum Genet 2000;66:187-195.
- 26. Takeda M, Kikuchi M, Ubalee R, Na-Bangchang K, Ruangweerayut R, Shibahara S, Imai S, Hirayama K. Microsatellite polymorphism in the heme oxygenease-1 gene promoter is associated with susceptibility to cerebral malaria in Myanmar. Jpn J Infect Dis 2005;58:268-271.
- 27. Zhuang H, Pin S, Li X, Doré S. Regulation of heme oxygenase expression by cyclopentenone prostaglandins. Exp Biol Med (Maywood) 2003;228:499-505.
- 28. Kubata BK, Eguchi N, Urade Y, Yamashita K, Mitamura T, Tai K, Hayaishi O, Horii T. Plasmodium falciparum produces prostaglandins that are pyrogenic, somnogenic, and immunosuppressive substances in humans. J Exp Med 1998;188:1197-1202.
- 29. Eguchi N, Minami T, Shirafuji N, Kanaoka Y, Tanaka T, Nagata A, Yoshida N, Urade Y, Ito S, Hayashi O. Lack of tactile pain (allodynia) in lipocalin-type prostaglandin D synthase-deficient mice. Proc Natl Acad Sci USA 1999;96:726-730.
- 30. Urade Y, Hayaishi O. Biochemical, structural, genetic, physiological, and pathophysiological features of lipocalin-type prostaglandin D synthase. Biochim Biophys Acta 2000;1482:259-271.
- 31. Urade Y, Fujimoto N, Hayaishi O. Purification and characterization of rat brain prostaglandin synthetase. J Biol Chem 1985;260:12410-12415.
- 32. Boie Y, Sawyer N, Slipetz DM, Metters KM, Abramovitz M. Molecular cloning and characterization of the human prostanoid DP receptor. J Biol Chem 1995;270:18910-18916.
- 33. Hirai H, Tanaka K, Yoshie O, Ogawa K, Kenmotsu K, Takamori Y, Ichimasa M, Sugamura K, Nakamura M, Takano S, Nagata K. Prostaglandin D2 selectively induces chemotaxis in T helper type 2 cells, eosinophils, and basophils via seven-transmembrane receptor CRTH2. J Exp Med 2001;193:255-261.
- 34. Pettipher R, Hansel TT, Armer R. Antagonism of the prostaglandin D2 receptors DP1 and CRTH2 as an approach to treat allergic diseases. Nat Rev Drug Discov 2007;6:313-325.
- 35. Obata T, Nakano Y, Looareesuwan S, Ohtomo H, Aikawa M. Prostaglandin spectrum in falciparum malaria patients. International Congress Series 2002;1233:475-478.
- 36. Kuesap J, Li B, Satarug S, Takeda K, Numata I, Na-Bangchang K, Shibahara S. Prostaglandin D2 induces heme oxygenase-1 in human retinal pigment epithelial cells. Biochem Biophys Res Commun 2008;367:413-419.
- 37. Beare NA, Taylor TE, Harding SP, Lewallen S, Molyneux ME. Malarial retinopathy: a newly established diagnostic sign in severe malaria. Am J Trop Med Hyg 2006;75:790-797.
- 38. Yoshida T, Biro P, Cohen T, Müller RM, Shibahara S. Human heme oxygenase cDNA and induction of its mRNA by hemin. Eur J Biochem 1988;171:457-461.
- 39. McCoubrey WK Jr, Ewing JF, Maines MD. Human heme oxygenase-2: characterization and expression of a full-length cDNA and evidence suggesting that the two HO-2 transcripts may differ by choice of polyadenylation signal. Arch Biochem Biophys 1992;295:13-20.
- 40. Arcari P, Martinelli R, Salvatore F. The complete sequence of a full length cDNA for human liver glyceraldehyde-3-phosphate dehydrogenase: evidence for multiple mRNA species. Nucleic Acids Res 1984;12:9179-9189.
- 41. Alvarez-Maqueda M, El Bekay R, Alba G, Monteseirín J, Chacón P, Vega A, Martín-Nieto J, Bedoya FJ, Pintado E, Sobrino F. 15-deoxy-delta 12,14-prostaglandin J2 induces heme oxygenase-1 gene expression in a reactive oxygen species-dependent manner in human lymphocytes. J Biol Chem 2004;279:21929-21937.
- 42. Qin S, McLaughlin AP, De Vries GW. Protection of RPE cells from oxidative injury by 15-deoxy-delta12,14-prostaglandin J2 by augmenting GSH and activating MAPK. Invest Ophthalmol Vis Sci 2006;47:5098-5105.
- 43. Kacem K, Lacombe P, Seylaz J, Bonvento G. Structural organization of the perivascular astrocyte endfeet and their relationship with the endothelial glucose transporter: a confocal microscopy study. Glia 1998;23:1-10.
- 44. Debault LE, Cancilla PA. Glutamyl transpeptidase in isolated brain endothelial cells: induction by glial cells in vitro. Science 1979;207:653-655.
- 45. Tontsch U, Bauer HC. Glial cells and neurons induce blood-brain barrier related enzymes in cultured cerebral endothelial cells. Brain Res 1991;539:247-253.
- 46. Harry GJ, Billingsley M, Bruinink A, Campbell IL, Classen W, Dorman DC, Galli C, Ray D, Smith RA, Tilson HA. In vitro techniques for the assessment of neurotoxicity. Environ Health Perspect 1998;106:131-158.
- 47. Viviani B, Corsini E, Galli CL, Padovani A, Ciusani E, Marinovich M. Dying neural cells activate glia through the release of a protease product. Glia 2000;32:84-90.
- 48. Lanz R, Polster P, Brune K. Antipyretic analgesics inhibit prostaglandin release from astrocytes and macrophages similarly. Eur J Pharmacol 1986;130:105-109.
- 49. Tzeng SF, Hsiao HY, Mak OT. Prostaglandins and cyclooxygenases in glial cells during brain inflammation. Curr Drug Targets Inflamm Allergy 2005;4:335-340.
- 50. Estrada C, Bready JV, Berliner JA, Pardridge WM, Cancilla PA. Astrocyte growth stimulation by a soluble factor produced by cerebral endothelial cells in vitro. J Neuropathol Exp Neurol 1990;49:539-549.
- 51. Sperri PE, Grant MB, Gomez J, Vernadakis A. Endothelial cell conditioned media mediated regulation of glutamine synthase activity in glial cells. Dev Brain Res 1997;104:205-208.
- 52. White NJ, Looareesuwan S, Warrell DA, Warrell MJ, Bunnag D, Harinasuta T. Quinine pharmacokinetics and toxicity in cerebral and uncomplicated falciparum malaria. Am J Med 1982;73:564-572.
- 53. Brewer TG, Genovese RF, Newman DB, Li Q. Factors relating to neurotoxicity of artemisinin antimalarial drugs "listening to artemether". Med Trop 1998;58:22-27.
- 54. Hoffman SL. Artemether in severe malaria-still too many deaths. N Engl J Med 1996;335:124-126.
- 55. Miller LG, Panosian CB. Ataxia and slurred speech after artesunate treatment for falciparum malaria. N Engl J Med 1997;336:1328.
- 56. Wesche DL, Da Coster MA, Tortella FC, Brewer TG. Neurotoxicity of artemisinin analogs in vitro. Antimicrob Agents Chemother 1994;38:1813-1819.
- 57. Karbwang J, Tin T, Rimchala W, Sukontason K, Namsiripongpun V, Thanavibul A, Na-Bangchang K, Laothavorn P, Bunnag D, Harinasuta T. Comparison of artemether and quinine in the treatment of severe falciparum malaria in south-east Thailand. Trans R Soc Trop Med Hyg 1995;89:668-671.
- 58. Phillips-Howard PA, ter Kuile FO. CNS adverse events associated with antimalarial agents: fact or fiction? Drug Safety 1995;12:370-383.
- 59. Löscher W, Potschka H. Blood-brain barrier active efflux transporters: ATP-binding cassette gene family. NeuroRx 2005;2:86-98.
- 60. Barraud LS, Fernandez C, German FM, Gantier JC, Gimenez F, Farinotti R. MDR1A (ABCB1) deficient CF-1 mutant mice is susceptible to cerebral malaria induced by Plasmodium berghei anka. J Parasitol 2008;94:1139-1142.
- 61. Aarnoudse AL, van Schaik RH, Dieleman J, Molokhia M, van Riemsdijk MM, Ligthelm RJ, Overbosch D, van der Heiden IP, Stricker BH. MDR1 gene polymorphisms are associated with neuropsychiatric adverse effects of mefloquine. Clin Pharmacol Ther 2006;80:367-374.
- 62. Sisodiya SM, Martinian L, Scheffer GL, van der Valk P, Scheper RJ, Harding BN, Thom M. Vascular colocalization of P-glycoprotein, multidrug-resistance associated protein 1, breast cancer resistance protein and major vault protein in human epileptogenic pathologies. Neuropathol Appl Neurobiol 2006;32:51-63.
- 63. Combes V, Coltel N, Faille D, Wassmer SC, Grau GE. Cerebral malaria: role of microparticles and platelets in alterations of the blood-brain barrier. Int J Parasitol 2006;36:541-546.
- 64. Giao PT, de Vries PJ. Pharmacokinetic interactions of antimalarial agents. Clin Pharmacokinet 2001;40:343-373.
Fig. 1Effects of PGD2 and 15dPGJ2 on HO-1 and HO-2 proteins in CCF-STTG1 cells. CCF-STTG1 human astrocyte cells were treated with PGD2 (A) or 15d-PGJ2 (B) at final concentration 0.5, 1, 5, or 10 µM for the indicated time from 3 to 24 hr and then harvested for preparation of proteins. Shown are the Western blots used for HO-1 and HO-2 protein. Each lane contained 30 µg proteins prepared from CCF-STTG1 cells. A bottom panel shows β-actin as an internal control. The data shown are from 1 of 2 independent experiments.
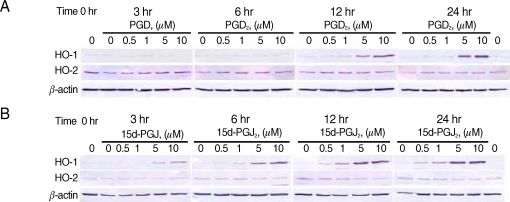
Fig. 2Expression of HO-1 mRNA by PGD2 and 15dPGJ2 in CCF-STTG1 astrocyte cells. CCF-STTG1 cells were treated with PGD2 or 15d-PGJ2 at final concentration 0.5, 1, 5, or 10 µM for the indicated time from 3 to 24 hr and then harvested for RNA preparation. cDNA were prepared for RT-PCR using Platinum® SYBR® Green qPCR SuperMix-UDG cocktail (Invitrogen). Shown are representative of the relative HO-1 mRNA and HO-2 mRNA expression of PGD2 (A, B) or 15d-PGJ2 (C, D) induction. The data were obtained by dividing the intensity value for each sample with 0-hr untreated control cells, which reflected a basal expression level.