Abstract
Bartonella species are vector-borne pathogens that infect a wide range of hosts, including humans. Although several Bartonella species have been identified in rodents and arthropods in Korea, information on Bartonella detection in ticks removed from human patients remains limited. This study investigated the presence of Bartonella species DNA in human-biting ticks collected in Korea and screened for other tick-associated bacteria, including Coxiella endosymbiont. From January to December 2018, 35 ticks were removed from 29 tick-bitten patients in Jeollanam-do and Gwangju, Korea. Ticks were identified morphologically and molecularly by 16S rRNA gene PCR. The presence of Bartonella species was assessed using nested PCR targeting the 16S-23S internal transcribed spacer (ITS) region. The ticks were identified as Haemaphysalis longicornis (17/35, 48.6%), Amblyomma testudinarium (14/35, 40.0%), and Ixodes nipponensis (4/35, 11.4%). Two H. longicornis ticks tested positive for Bartonella species. Sequencing revealed 99.5% identity with B. bacilliformis isolate GJRITS124 in one tick and 98.9% identity with B. taylorii isolate 190731_HC2 in the other, both previously identified in Apodemus agrarius rodents in Korea. One B. bacilliformis-positive tick was also positive for Coxiella spp., and sequence analysis indicated a Coxiella endosymbiont showing 100.0% identity with the Coxiella-like endosymbiont strain 580. Phylogenetic analysis supported these findings; however, bacterial cultures from PCR-positive tick lysates were negative. This study provides baseline evidence of B. bacilliformis and B. taylorii, as well as Coxiella endosymbiont DNA in human-biting H. longicornis ticks in Korea and highlights the need for continued surveillance.
-
Key words: Ticks, Bartonella, Coxiella, tick-borne infections, polymerase chain reaction
Ectoparasites, such as ticks, are prominent vectors of numerous infectious diseases, including those caused by bacteria, viruses, and protozoans. Ticks are haematophagous arthropods, and their ability to acquire and maintain pathogens is influenced by complex tick-pathogen interactions and vector competence-related mechanisms [
1]. However, for
Bartonella species, the role of ticks as vectors remains uncertain and has not been conclusively demonstrated.
Bartonella species are zoonotic bacteria present in various mammals, including rodents, cats, ruminants, and humans [
2]. Although several studies have suggested possible tick-associated transmission, their role as definitive vectors remains controversial [
3]. Molecular surveys have detected
Bartonella DNA in multiple tick genera, including
Haemaphysalis and
Ixodes across different geographic regions [
4,
5]. However, direct transmission of
Bartonella species from ticks to mammalian hosts has not been conclusively demonstrated [
3]. Among
Bartonella species,
B. bacilliformis is a human-pathogenic organism typically associated with sandfly transmission, whereas
B. taylorii is mainly maintained in rodent-flea cycles [
6,
7]. The involvement of ticks in the ecology of these species has not been established.
In East Asia,
Bartonella DNA has been detected in ticks, including
H. longicornis, collected in China, indicating that this tick species can harbor
Bartonella in the region [
5]. In Korea, multiple
Bartonella species, including
B. grahamii,
B. henselae,
B. taylorii,
B. tribocorum, and
B. phoceensis, have been identified in
Apodemus agrarius rodents [
8]. Additionally,
H. longicornis, a dominant tick species in East Asia, is known to transmit several pathogens, including
Rickettsia,
Anaplasma, and severe fever with thrombocytopenia syndrome virus [
5,
9-
11]. Given the increasing reports of
Bartonella species in ticks globally, it is crucial to investigate their potential involvement in the transmission of
Bartonella species. Understanding the interaction between
Bartonella species and their potential tick vectors is critical for evaluating the epidemiological impact of tick-borne
Bartonella infections. Therefore, this study investigated the molecular detection of
Bartonella species DNA in ticks removed from human patients in Korea and performed additional screening PCR assays for other tick-associated pathogens, including
Rickettsia spp.,
Orientia tsutsugamushi,
Anaplasma spp., and
Coxiella spp., to evaluate their occurrence at the human–tick interface.
Between January and December 2018, ticks removed from humans in Jeollanam-do and Gwangju, Korea, were collected during routine clinical care for tick-bitten patients at Chosun University Hospital, and sampling was based on clinical presentation rather than a predefined geographic or temporal survey design. The study was approved by the Ethics in Human Research Committee of Chosun University Hospital under an institutional review board that approves all experiments involving ticks and tick-bitten humans (No. CHOSUN 2013-10-001-018). Ticks were morphologically identified to the species and developmental stage (adult female, adult male, nymph, or larva) using standard taxonomic parameters and morphological features under a microscope [
12]. Each identified tick was individually processed and subjected to molecular identification using conventional PCR (C-PCR) targeting mitochondrial
16S rRNA [
13].
Ticks at all developmental stages were washed in 70% ethanol and rinsed twice with sterile phosphate-buffered saline. Each tick was transferred to a hard tissue grinding MK28 tube (Bertin Technology) containing 800 μl of phosphate-buffered saline with 1× PC/SM (penicillin and streptomycin), ground using a FastPrep-24 Classic instrument (MP Biomedicals), and then stored at -80°C until DNA extraction. For DNA extraction, 150 µl of the homogenized tick lysate was mixed with 150 µl ATL buffer and 20 µl proteinase K and incubated at 56°C overnight for lysis; genomic DNA was extracted using the QIAamp Tissue & Blood Mini Kit (Qiagen) according to the manufacturer’s instructions.
PCR amplification was performed using target-specific primers and AmpliTaq Gold 360 Master Mix (Applied Biosystems) on an AB thermal cycler (Applied Biosystems). The 16S-23S internal transcribed spacer (
ITS) region was targeted for detection of
Bartonella species [
8], and PCR reactions included positive controls (
B. henselae DNA) and negative controls (molecular-grade water). Additional screening PCR assays were performed for
Rickettsia spp.,
Orientia tsutsugamushi,
Anaplasma spp., and
Coxiella spp. using previously described primers and conditions (
Supplementary Table S1) [
8,
13-
17]. For nested PCR, the reaction mixture was identical to that used for C-PCR, except that the first PCR product was used as template DNA with nested PCR primers, and the screening results are presented in
Table 1.
The amplified PCR products were purified using the QIAquick PCR Purification Kit (Qiagen) and sequenced with PCR primers (Solgent). Obtained sequences were compared with GenBank database entries using the Basic Local Alignment Search Tool (BLAST) provided by the National Center for Biotechnology Information and aligned using Lasergene version 8 (DNASTAR). Phylogenetic trees were constructed by the neighbor-joining method based on ClustalW alignments using the MegAlign program (DNASTAR). Bootstrap analysis (1,000 replicates) was performed using the Kimura 2-parameter model. Pairwise alignments were performed with an open-gap penalty of 10 and a gap extension penalty of 0.5. Bacterial culture of
Bartonella species PCR-positive tick lysates was performed on heart infusion agar blood plates using 200 μl of the homogenized tick lysate [
18].
All ticks examined in this study were fed or partially fed ticks that had been removed from human patients during clinical visits. From 29 tick-bitten patients, a total of 35 ticks were obtained, of which 19 ticks (54.3%) were adults, including 16 (45.7%) females and 3 males (8.6%). Thirteen ticks (37.1%) were nymphs, and 3 (8.6%) were larvae based on morphological examination using a microscope for tick identification. The ticks were identified as
H. longicornis (17 ticks, 48.6%; 12 adult females, 2 nymphs, and 3 larvae),
Amblyomma testudinarium (14 ticks, 40.0%; 3 adult males and 11 nymphs), and
Ixodes nipponensis (4 ticks, 11.4%; 4 adult females), as described in
Table 2. Tick identification using 16S rDNA C-PCR and DNA sequencing yielded the same results as microscopic examination (
Table 2).
A total of 35 ticks were examined for the detection of
Bartonella species using
Bartonella-specific 16S-23S
ITS nested PCR. Among these, 2 ticks (2 out of 35, 5.7%) tested positive for
Bartonella species. Both
Bartonella-positive ticks were identified as
H. longicornis (
Table 1;
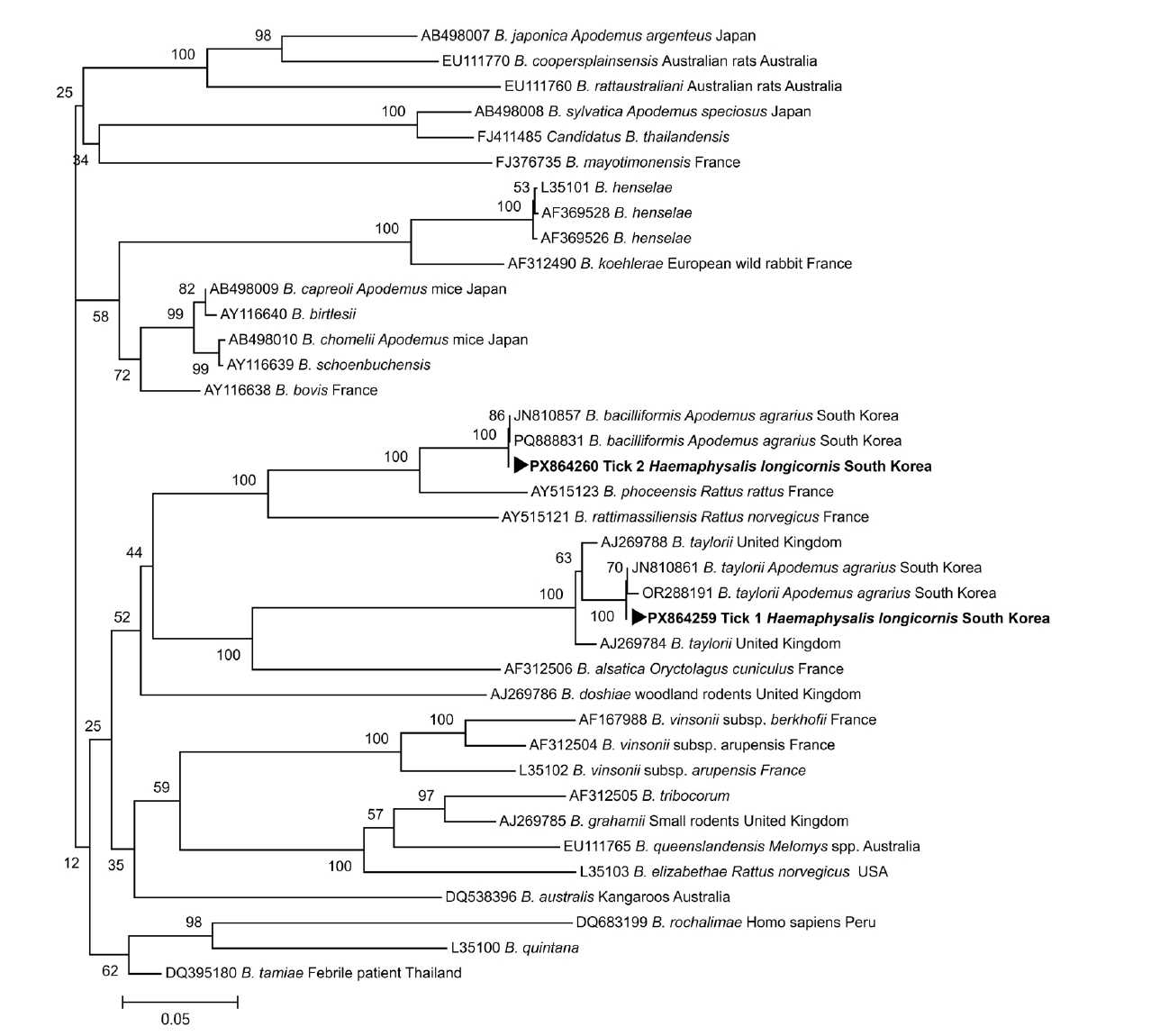
Supplementary Fig. S1). The positive PCR products were sequenced, and the resulting sequences were aligned with reference sequences obtained from the GenBank database to identify known sequences with a high degree of similarity using ClustalW. The DNA sequence analysis revealed the presence of
B. taylorii in tick 1 and
B. bacilliformis in tick 2. Homology testing showed that the
B. taylorii-positive tick 1 exhibited 98.9% identity with
B. taylorii isolate 190731_HC2 (GenBank accession No. OR288191.1), and the
B. bacilliformis-positive tick 2 exhibited 99.5% identity with
B. bacilliformis isolate GJR
ITS124 (GenBank accession No. PQ888831.1), both previously identified in
A. agrarius rodents in Korea. A phylogenetic tree was constructed based on the alignment of 16S–23S
ITS sequences obtained from the 2
Bartonella-positive
H. longicornis ticks together with reference
Bartonella sequences retrieved from GenBank. In the phylogenetic analysis, the sequence from tick 1 clustered with
B. taylorii, whereas the sequence from tick 2 clustered with
B. bacilliformis (
Fig. 1), supporting the species identification based on sequence similarity. The 16S–23S
ITS nested PCR product sequences generated in this study were deposited in the GenBank database under accession numbers PX864259 (
B. taylorii, tick 1) and PX864260 (
B. bacilliformis, tick 2).
No DNA of
Rickettsia spp.,
O. tsutsugamushi, or
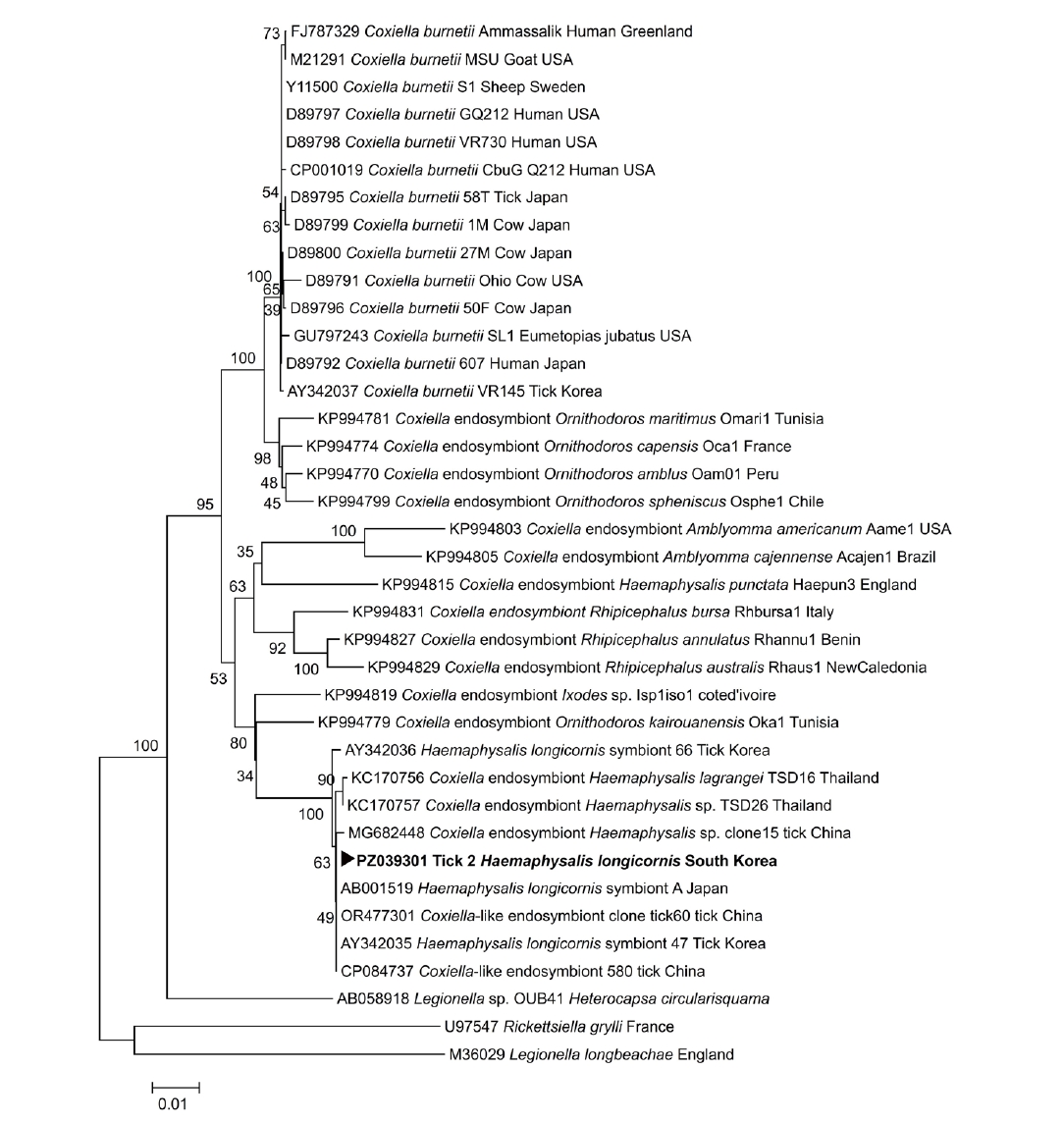
Anaplasma spp. was detected in any of the examined ticks. One tick (tick 2) tested positive by PCR for
Coxiella spp.; however, sequence analysis indicated that this signal corresponded to a
Coxiella endosymbiont rather than
Coxiella burnetii (
Table 1). Homology analysis demonstrated that the
Coxiella endosymbiont–positive tick 2 exhibited 100.0% identity with the
Coxiella-like endosymbiont strain 580 chromosome (GenBank accession No. CP084737.1). The partial
16S rRNA gene sequence of the
Coxiella endosymbiont detected in
H. longicornis tick 2 was deposited in the GenBank database under accession number PZ039301 (
Coxiella endosymbiont, tick 2). A phylogenetic tree based on the partial
16S rRNA sequence obtained from the
Coxiella endosymbiont-positive tick 2 was constructed with reference sequences from GenBank, and the sequence clustered with
Coxiella endosymbionts (
Fig. 2). No viable
Bartonella colonies were obtained from PCR-positive tick lysates, indicating the difficulty of isolating viable
Bartonella organisms from tick vectors under the culture conditions used.
Detection of
B. bacilliformis and
B. taylorii DNA and
Coxiella endosymbiont DNA in human-biting
H. longicornis ticks demonstrates the presence of tick-associated bacterial DNA at the human-tick interface in Korea. However, molecular detection alone does not demonstrate transmission by ticks. Previous studies have also detected
Bartonella DNA in various tick genera worldwide [
4,
5], indicating that ticks may harbor the organism under natural conditions [
4].
Phylogenetic analysis showed that the Bartonella sequences detected in human-biting ticks were closely related to strains previously reported from A. agrarius rodents in Korea, showing 99.5% identity for B. bacilliformis and 98.9% identity for B. taylorii. These findings suggest that the Bartonella DNA detected in ticks removed from human patients showed sequence similarity to strains previously detected in rodents in Korea. However, the ecological or epidemiological relationships among rodents, ticks, and humans cannot be determined based solely on the molecular data obtained in this study.
The 2 Bartonella-positive H. longicornis ticks were collected from different patients. One tick (tick 1) positive for B. taylorii was removed from a patient (2018-Lee01) who presented solely for tick removal without fever or other systemic symptoms suggestive of bartonellosis. The other tick (tick 2), which tested positive for B. bacilliformis, was removed from a different patient (2018-505) who presented with fever; however, no clinical features characteristic of bartonellosis, such as Oroya fever, verruga peruana, endocarditis, or lymphadenopathy, were identified during clinical evaluation. No laboratory or clinical evidence of confirmed human Bartonella infection was documented in either patient.
One possible explanation for this sequence similarity is that the detected
Bartonella DNA may have originated from the blood meal of infected rodent hosts, such as
A. agrarius, rather than reflecting active infection or transmission by the tick, as previous studies have emphasized that detection of
Bartonella DNA in ticks does not necessarily indicate vector competence [
3]. Consistent with this interpretation, bacterial cultures of PCR-positive tick lysates were negative in this study, further illustrating the difficulty of isolating viable
Bartonella organisms and highlighting the limitations of current culture-based approaches.
This study has several limitations, including the relatively small number of ticks examined, the restriction of sampling to a single year, and the clinical-based consecutive collection of ticks from patients at a single hospital, which limits epidemiological interpretation. Nevertheless, previous studies in Korea have reported the presence of
Bartonella DNA in various tick species and rodents, and human infections have also been reported, supporting the zoonotic relevance of these organisms [
4,
19]. Similar molecular findings from other regions, including the detection of
B. taylorii in rodent species such as
A. flavicollis,
A. sylvaticus, and
Clethrionomys glareolus in Slovenia, indicate that these pathogens occur across diverse geographic settings and host species [
20]. Future studies incorporating quantitative PCR to estimate bacterial load, multilocus sequence typing, or metagenomic approaches would help further validate species identification and provide a more comprehensive understanding of
Bartonella diversity detected in human-biting ticks.
In conclusion, this study provides baseline evidence for the presence of B. bacilliformis and B. taylorii, as well as Coxiella endosymbiont DNA in H. longicornis ticks removed from human patients in Korea. The findings are limited to molecular detection in a small number of fed ticks and do not demonstrate transmission by ticks. These results provide baseline detection data for tick-associated bacterial DNA, including Bartonella and Coxiella endosymbiont, in human-biting ticks in Korea and support further studies to clarify the ecological relevance of these findings in human exposure settings.
Notes
-
Data availability
Data and materials are available upon request to the corresponding author.
-
Author contributions
Conceptualization: Kim DM. Data curation: Kim CM, Lee YM. Formal analysis: Kim CM, Panchali MJL. Funding acquisition: Kim DM. Investigation: Kim CM, Panchali MJL, Lee YM. Methodology: Lee YM. Project administration: Kim DM. Resources: Lee YM, Yun NR. Supervision: Kim DM, Yun NR. Validation: Kim CM, Panchali MJL. Visualization: Lee YM, Yun NR. Writing - original draft: Kim CM, Panchali MJL. Writing - review & editing: Kim DM, Kim CM, Panchali MJL.
-
Conflict of interest
The authors have no conflicts of interest to declare.
-
Funding
This study was supported by a research fund from Chosun University Hospital (2025).
Supplementary information
Fig. 1.Phylogenetic tree constructed based on partial 16S–23S internal transcribed spacer (ITS) sequences of the Bartonella spp. detected in 2 Haemaphysalis longicornis ticks (tick 1 and tick 2) removed from human patients in Korea (▶), together with reference Bartonella sequences retrieved from the GenBank database. The phylogenetic tree was constructed using the neighbor-joining method. Bootstrap values based on 1,000 replicates are shown at the branch nodes.The scale bar represents 0.05 nucleotide substitutions per site. GenBank accession numbers are indicated for all reference sequences. Host and country information for the reference sequences are provided. For some reference sequences retrieved from GenBank, information on the host and/or country of origin was not available in the original database records.
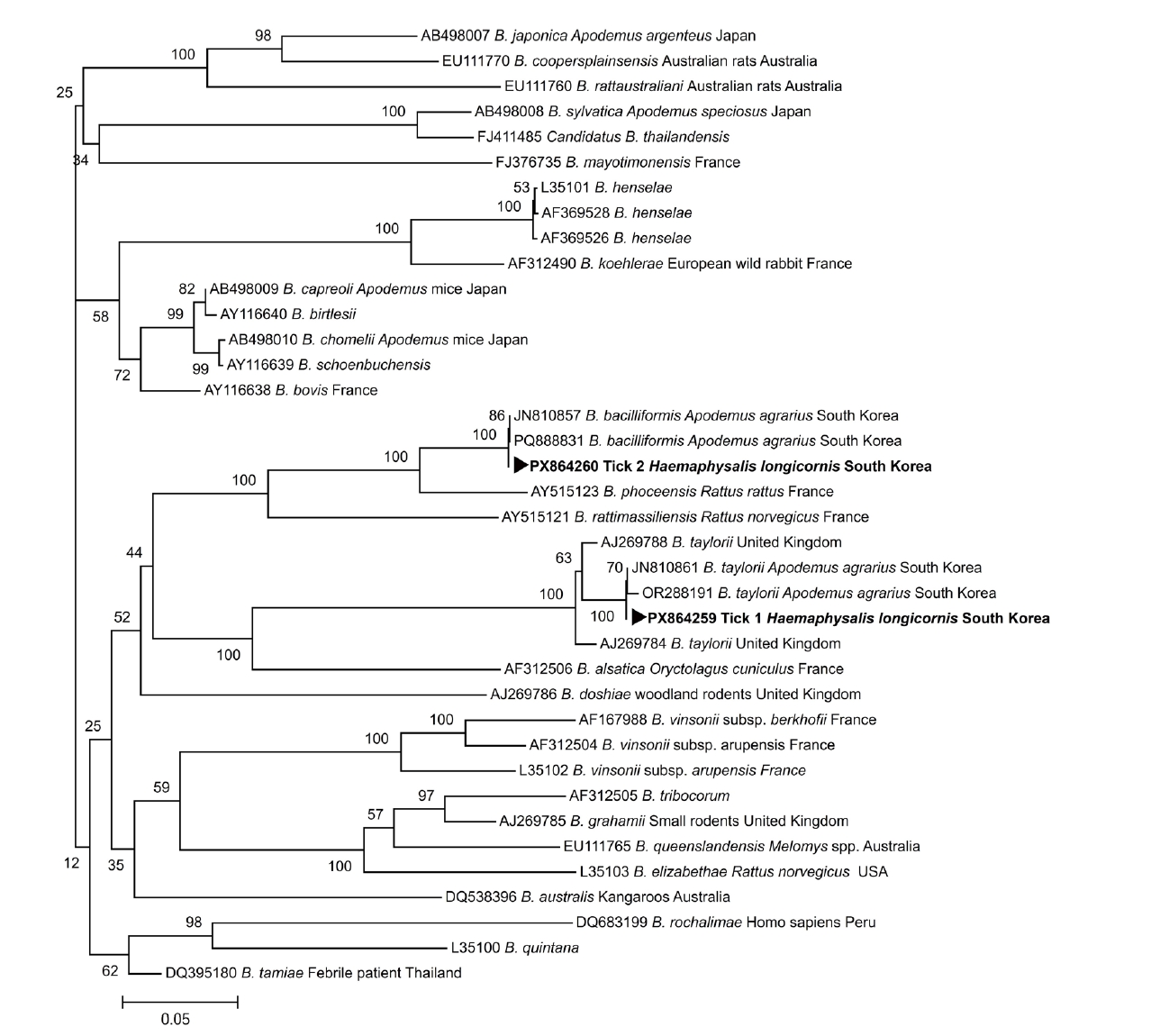
Fig. 2.Phylogenetic tree constructed based on partial 16S rRNA gene sequences of the Coxiella endosymbiont detected in a Haemaphysalis longicornis tick (tick 2) removed from human patients in Korea (▶), together with reference sequences of Coxiella burnetii and Coxiella endosymbionts retrieved from the GenBank database. The phylogenetic tree was constructed using the neighbor-joining method. Bootstrap values based on 1,000 replicates are shown at the branch nodes. The scale bar represents 0.01 nucleotide substitutions per site. GenBank accession numbers are indicated for all reference sequences.
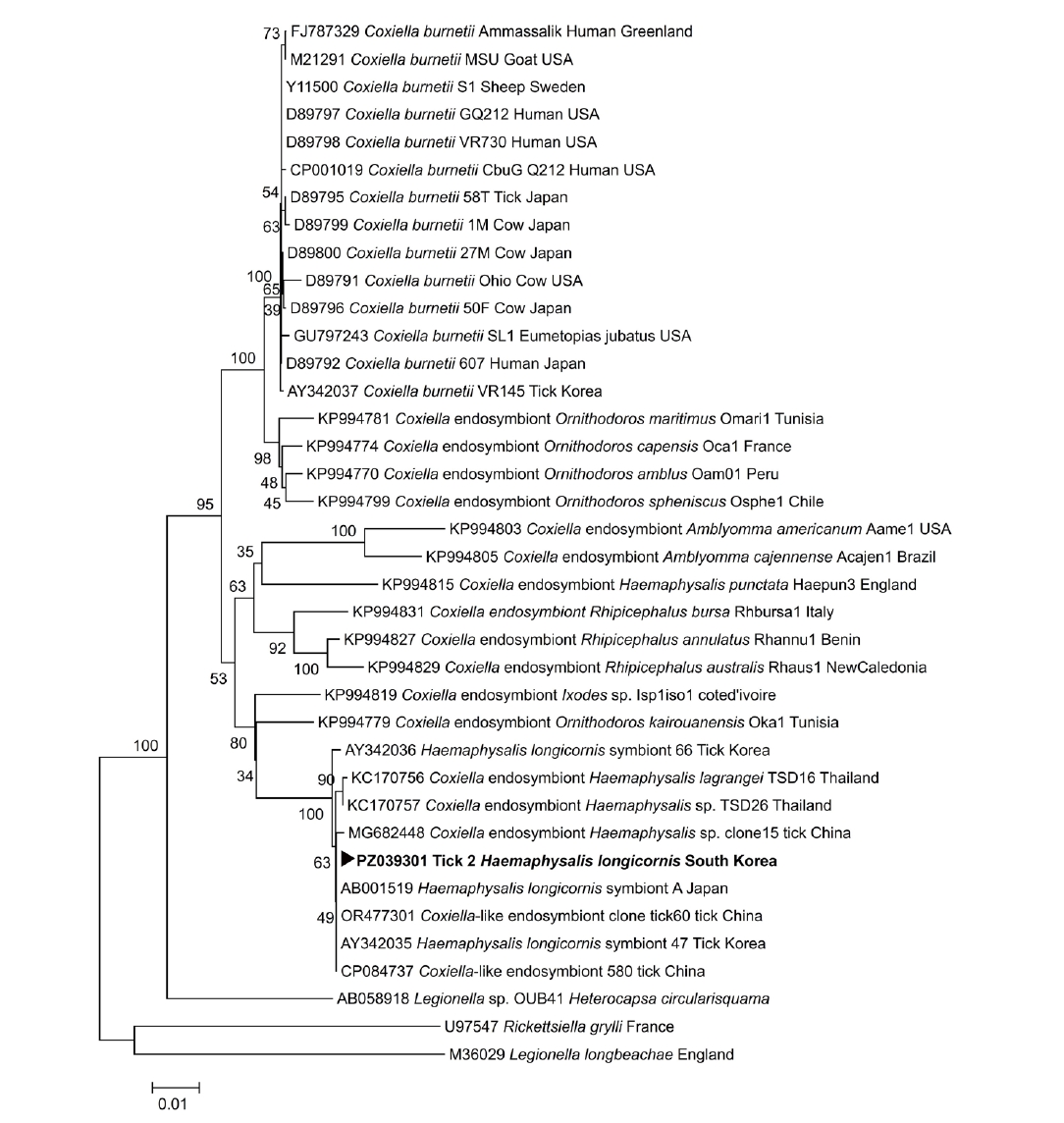
Table 1.Identification of ticks and detection of tick-borne pathogens in ticks collected from human patients
Table 1.
|
Tick No. |
Tick species |
Tick developmental stage |
Rickettsia spp. |
Orientia tsutsugamushi
|
Anaplasma spp. |
Coxiella spp. |
Bartonella spp. |
|
ompA
|
56 kDa
|
groEL ankA
|
16S rRNA IS1111
|
ITS
|
|
Tick 1 |
Haemaphysalis longicornis
|
Adult female |
- |
- |
- |
- |
- |
- |
Bartonella taylorii
|
|
Tick 2 |
H. longicornis
|
Adult female |
- |
- |
- |
- |
Coxiella endosymbiont |
- |
Bartonella bacilliformis
|
Table 2.16S rDNA-targeting conventional PCR and morphological identification of ticks collected from patients
Table 2.
|
Development stage/tick species |
Haemaphysalis longicornis
|
Amblyomma testudinarium
|
Ixodes nipponensis
|
Species total |
|
Adult female |
12 (70.6) |
0 (0) |
4 (100) |
16 (45.7) |
|
Adult male |
0 (0) |
3 (21.4) |
0 (0) |
3 (8.6) |
|
Nymph |
2 (11.8) |
11 (78.6) |
0 (0) |
13 (37.1) |
|
Larva |
3 (17.6) |
0 (0) |
0 (0) |
3 (8.6) |
|
Stage total |
17 (48.6) |
14 (40.0) |
4 (11.4) |
35 (100) |
References
- 1. de la Fuente J, Antunes S, Bonnet S, et al. Tick-pathogen interactions and vector competence: identification of molecular drivers for tick-borne diseases. Front Cell Infect Microbiol 2017;7:114. https://doi.org/10.3389/fcimb.2017.00114
- 2. Deng H, Pang Q, Zhao B, Vayssier-Taussat M. Molecular mechanisms of Bartonella and mammalian erythrocyte interactions: a review. Front Cell Infect Microbiol 2018;8:431. https://doi.org/10.3389/fcimb.2018.00431
- 3. Angelakis E, Billeter SA, Breitschwerdt EB, Chomel BB, Raoult D. Potential for tick-borne bartonelloses. Emerg Infect Dis 2010;16:385-91. https://doi.org/10.3201/eid1603.081685
- 4. Kim CM, Kim JY, Yi YH, et al. Detection of Bartonella species from ticks, mites and small mammals in Korea. J Vet Sci 2005;6:327-34. https://doi.org/10.4142/jvs.2005.6.4.327
- 5. Sun J, Liu Q, Lu L, et al. Coinfection with four genera of bacteria (Borrelia, Bartonella, Anaplasma, and Ehrlichia) in Haemaphysalis longicornis and Ixodes sinensis ticks from China. Vector Borne Zoonotic Dis 2008;8:791-5. https://doi.org/10.1089/vbz.2008.0005
- 6. Bai Y, Kosoy MY, Boonmar S, et al. Enrichment culture and molecular identification of diverse Bartonella species in stray dogs. Vet Microbiol 2010;146:314-9. https://doi.org/10.1016/j.vetmic.2010.05.017
- 7. Marié JL, Fournier PE, Rolain JM, et al. Molecular detection of Bartonella quintana, B. elizabethae, B. koehlerae, B. doshiae, B. taylorii, and Rickettsia felis in rodent fleas collected in Kabul, Afghanistan. Am J Trop Med Hyg 2006;74:436-9. https://doi.org/10.4269/ajtmh.2006.74.436
- 8. Ko S, Kang JG, Kim HC, et al. Prevalence, isolation and molecular characterization of Bartonella species in Republic of Korea. Transbound Emerg Dis 2016;63:56-67. https://doi.org/10.1111/tbed.12217
- 9. Kim CM, Kim MS, Park MS, Park JH, Chae JS. Identification of Ehrlichia chaffeensis, Anaplasma phagocytophilum, and A. bovis in Haemaphysalis longicornis and Ixodes persulcatus ticks from Korea. Vector Borne Zoonotic Dis 2003;3:17-26. https://doi.org/10.1089/153036603765627424
- 10. Lee J, Moon K, Kim M, et al. Seasonal distribution of Haemaphysalis longicornis (Acari: Ixodidae) and detection of SFTS virus in Gyeongbuk Province, Republic of Korea, 2018. Acta Trop 2021;221:106012. https://doi.org/10.1016/j.actatropica.2021.106012
- 11. Stanley HM, Ford SL, Snellgrove AN, et al. The ability of the invasive Asian longhorned tick Haemaphysalis longicornis (Acari: Ixodidae) to acquire and transmit Rickettsia rickettsii (Rickettsiales: Rickettsiaceae), the agent of Rocky Mountain spotted fever, under laboratory conditions. J Med Entomol 2020;57:1635-9. https://doi.org/10.1093/jme/tjaa076
- 12. Yamaguti N, Tipton VJ, Keegan HL, Toshioka S. Ticks of Japan, Korea, and the Ryukyu Islands. Brigh Young Univ Sci Bull Biol Ser 1971;15:Article 1.
- 13. Tian Z, Liu G, Xie J, et al. Discrimination between Haemaphysalis longicornis and H.s qinghaiensis based on the partial 16S rDNA and the second internal transcribed spacer (ITS-2). Exp Appl Acarol 2011;54:165-72. https://doi.org/10.1007/s10493-010-9423-3
- 14. Bang MS, Kim CM, Park JW, et al. Prevalence of Orientia tsutsugamushi, Anaplasma phagocytophilum and Leptospira interrogans in striped field mice in Gwangju, Republic of Korea. PLoS One 2019;14:e0215526. https://doi.org/10.1371/journal.pone.0215526
- 15. Fenollar F, Fournier PE, Raoult D. Molecular detection of Coxiella burnetii in the sera of patients with Q fever endocarditis or vascular infection. J Clin Microbiol 2004;42:4919-24. https://doi.org/10.1128/JCM.42.11.4919-4924.2004
- 16. Regnery RL, Spruill CL, Plikaytis BD. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J Bacteriol 1991;173:1576-89. https://doi.org/10.1128/jb.173.5.1576-1589.1991
- 17. Roux V, Fournier PE, Raoult D. Differentiation of spotted fever group rickettsiae by sequencing and analysis of restriction fragment length polymorphism of PCR-amplified DNA of the gene encoding the protein rOmpA. J Clin Microbiol 1996;34:2058-65. https://doi.org/10.1128/jcm.34.9.2058-2065.1996
- 18. Minnick MF, Smitherman LS, Samuels DS. Mitogenic effect of Bartonella bacilliformis on human vascular endothelial cells and involvement of GroEL. Infect Immun 2003;71:6933-42. https://doi.org/10.1128/IAI.71.12.6933-6942.2003
- 19. Durey A, Kwon HY, Im JH, et al. Bartonella henselae infection presenting with a picture of adult-onset Still's disease. Int J Infect Dis 2016;46:61-3. https://doi.org/10.1016/j.ijid.2016.03.014
- 20. Knap N, Duh D, Birtles R, et al. Molecular detection of Bartonella species infecting rodents in Slovenia. FEMS Immunol Med Microbiol 2007;50:45-50. https://doi.org/10.1111/j.1574-695X.2007.00226.x